This chapter focuses on the structure, biosynthesis, and general biology of proteoglycans. Topics include a description of the major families of proteoglycans, their characteristic polysaccharide chains (glycosaminoglycans), biosynthetic pathways, and general concepts about proteoglycan function. Proteoglycans, like other glycoconjugates, have many essential roles in biology.
HISTORICAL PERSPECTIVE
The study of proteoglycans dates back to the beginning of the 20th century with investigations of “chondromucoid” from cartilage and anticoagulant preparations from liver (heparin). From 1930 to 1960, great strides were made in analyzing the chemistry of the polysaccharides of these preparations (also known as “mucopolysaccharides”), yielding the structure of hyaluronan (Chapter 16), dermatan sulfate (DS), keratan sulfate (KS), different isomeric forms of chondroitin sulfate (CS), heparin, and heparan sulfate (HS). Together, these polysaccharides came to be known as glycosaminoglycans (sometimes abbreviated as GAGs) to indicate the presence of amino sugars and other sugars in a polymeric form. Subsequent studies provided insights into the linkage of the chains to proteoglycan core proteins. These structural studies paved the way for biosynthetic studies.
The 1970s marked a turning point in the field, when improved isolation and chromatographic procedures were developed for the purification and analysis of tissue proteoglycans and glycosaminoglycans. Density-gradient ultracentrifugation allowed separation of the large aggregating proteoglycans from cartilage, revealing a complex of proteoglycan, hyaluronan, and link protein. Also during this period, it was realized that the production of proteoglycans was a general property of animal cells and that proteoglycans and glycosaminoglycans were present on the cell surface, inside the cell, and in the extracellular matrix (ECM). This observation led to a rapid expansion of the field and the eventual appreciation of proteoglycan function in cell adhesion, signaling, and other biological activities (Chapter 38). Today, studies with animal cell mutants (Chapter 49) as well as experiments using gene knockout and silencing techniques in a variety of model organisms are aimed at extending our understanding of the role of proteoglycans in development and physiology (Chapters 25–27) and human diseases (Chapters 41–47). The application of a variety of newly developed analytical tools, including mass spectrometry (Chapter 50) and glycan arrays (Chapter 48), are leading to a better understanding of proteoglycan structure and function.
PROTEOGLYCAN AND GLYCOSAMINOGLYCAN COMPOSITION
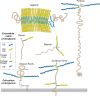
Proteoglycans consist of a “core” protein and one or more covalently attached glycosaminoglycan chains (Figure ). Glycosaminoglycans are linear polysaccharides that can be broken down chemically or enzymatically into disaccharides, each consisting of an amino sugar (glucosamine [GlcN] that is N-acetylated [GlcNAc] or N-sulfated [GlcNS] or N-acetylgalactosamine [GalNAc]) and a uronic acid (glucuronic acid [GlcA] or iduronic acid [IdoA]) or galactose (Gal). Figure depicts schematic short segments of glycosaminoglycans and their characteristic features. Hyaluronan does not occur covalently linked to a protein core but instead interacts noncovalently with some proteoglycans via hyaluronan-binding motifs (Chapter 16). Generally, invertebrates produce the same types of glycosaminoglycans as vertebrates, except that hyaluronan is not present and the chondroitin chains are predominantly, although not exclusively, nonsulfated. Most proteoglycans also contain N- and O-glycans typically found on glycoproteins (see Chapters 9 and 10). The glycosaminoglycan chains are much larger than these other types of glycans (e.g., a 20-kDa glycosaminoglycan chain contains approximately 80 sugar residues, whereas a typical biantennary N-glycan contains 10 to 12 residues). Keratan sulfate is a sulfated poly-N-acetyllactosamine chain present on a limited number of proteins as an N-linked or O-linked chain. The composition of the glycosaminoglycan chains, the structure of the protein cores, and the distribution of the proteoglycan all determine the biological activities associated with proteoglycans.
Proteoglycans consist of a protein core (brown) and one or more covalently attached glycosaminoglycan chains (dark blue, HS, heparan sulfate; yellow, CS/DS, chondroitin sulfate/dermatan sulfate; light blue, KS, keratan sulfate). Membrane proteoglycans (more...)
Glycosaminoglycans consist of alternating N-acetylated (GlcNAc or GalNAc) or N-sulfated (GlcNS) glucosamine and either a uronic acid (GlcA or IdoA) or galactose (Gal). Hyaluronan lacks sulfate groups, but the rest of the glycosaminoglycans contain sulfates (more...)
PROTEOGLYCANS ARE DIVERSE IN STRUCTURE AND FUNCTION
Virtually all mammalian cells produce proteoglycans and secrete them into the ECM, insert them into the plasma membrane, or store them in secretory granules. The ECM, an essential component of all multicellular animals, determines the physical characteristics of tissues and many of the biological properties of the cells embedded in it. The major components of the ECM are fibrillar proteins that provide tensile strength and elasticity (e.g., various collagens and elastins), adhesive glycoproteins (e.g., fibronectin, laminins, and tenascins), and proteoglycans that interact with other ECM components to promote ECM assembly, govern its physical properties, and serve as a reservoir of biologically active small proteins such as growth factors. A single cell type can express multiple proteoglycans. Vascular endothelial cells, for example, synthesize several different cell-surface proteoglycans, secretory granule proteoglycans, as well as several ECM proteoglycans.
Compared with the hundreds, perhaps thousands, of glycoproteins that carry N- and O-linked glycans, to date, relatively few proteins have been identified that carry glycosaminoglycans (less than 50), but the application of new glycoproteomic approaches has already led to the discovery of several new proteoglycans. Tremendous structural variation of proteoglycans exists due to a number of factors. First, many proteoglycans can be substituted with one or more types of glycosaminoglycan chain; for example, glypicans contain heparan sulfate, whereas syndecan-1 contains both heparan sulfate and chondroitin sulfate chains. Some proteoglycans contain only one glycosaminoglycan chain (e.g., decorin), whereas others have more than 100 chains (e.g., aggrecan). Another source of variability lies in the stoichiometry of glycosaminoglycan chain substitution. For example, syndecan-1 has five attachment sites for glycosaminoglycans, but not all of the sites are used equally. Other proteoglycans can be “part-time”—that is, they may exist with or without a glycosaminoglycan chain or with only a truncated oligosaccharide. A given proteoglycan present in different cell types often shows differences in the number of glycosaminoglycan chains, their lengths, and the arrangement of sulfated residues along the chains. Thus, a preparation of any one proteoglycan (defined by its core protein) represents a diverse population of molecules, each potentially representing a unique structural entity. These characteristics, typical of all proteoglycans, create enormous diversity and potential biological variation in activity.
Mammalian Proteoglycans—Form and Function
The major classes of proteoglycans can be classified by their distribution, homologies, and function. Table provides an overview of many of the well-known and characterized proteoglycans.
Diversity among known vertebrate proteoglycans
The aggrecan family of ECM proteoglycans (also known as lecticans) consists of aggrecan, versican, brevican, and neurocan. In all four members, the protein moiety contains an amino-terminal domain capable of binding hyaluronan, a central region that contains covalently bound chondroitin sulfate chains, and a carboxy-terminal domain containing a C-type lectin domain (Chapter 34). Aggrecan is the best-studied member of this family, because it represents the major proteoglycan in cartilage where it forms a stable matrix capable of withstanding compressive forces by water desorption and resorption. Versican, which is produced predominantly by connective tissue cells, undergoes alternative splicing events that generate a family of proteins. Neurocan is expressed in the late embryonic central nervous system (CNS) and can inhibit neurite outgrowth. Brevican is expressed in the terminally differentiated CNS, particularly in perineuronal nets.
The small leucine-rich proteoglycans (SLRPs) contain leucine-rich repeats flanked by cysteines in their central domain. At least nine members of this family are known and some carry chondroitin sulfate, dermatan sulfate, or keratan sulfate chains. These proteoglycans help to stabilize and organize collagen fibers but have other roles in innate immunity and regulation of growth factor signaling.
The SLRPs and aggrecan family of proteoglycans appear to be unique to vertebrates. Caenorhabditis elegans and Drosophila melanogaster express other proteoglycans, suggesting that the core proteins have undergone enormous diversification during evolution, presumably to accommodate different needs of the organism. In contrast, the biosynthetic machinery for glycosaminoglycan assembly has been evolutionarily conserved, demonstrating conservation of function for the glycosaminoglycan chains.
Basement membranes are highly specialized thin layers of the ECM that lie flush against epithelial cells and surround muscle and fat cells. Major components are laminins, nidogens, and collagens, as well as three unrelated basement membrane proteoglycans—perlecan, agrin, and type XVIII collagen. These proteoglycans interact with other basement membrane components and cell-surface adhesion receptors but can also be important reservoirs of heparan sulfate–binding signaling factors.
The membrane-bound proteoglycans are diverse. The syndecan family consists of four members, each with a short hydrophobic domain that spans the membrane, linking the larger extracellular domain containing the glycosaminoglycan attachment sites to a smaller intracellular cytoplasmic domain. The syndecans are expressed in a tissue-specific manner and facilitate cellular interactions with a wide range of extracellular ligands, such as growth factors and matrix molecules. Because of their membrane-spanning properties, the syndecans can transmit signals from the extracellular environment to the intracellular cytoskeleton via their cytoplasmic tails. Syndecans are sensitive to proteolytic cleavage by matrix metalloproteases, resulting in shedding of the ectodomains bearing the glycosaminoglycan chains that retain potent biological activity (Chapter 38). C. elegans and D. melanogaster express only one syndecan (Chapters 25 and 26).
Glypicans carry only HS chains, which can bind a wide array of factors essential for development and morphogenesis. Six glypican family members exist in mammals, and only two are expressed in D. melanogaster and C. elegans. Each member of the glypican family of cell-surface proteoglycans has a glycosylphosphatidylinositol anchor attached at the carboxyl terminus, which embeds them in the outer leaflet of the plasma membrane (Chapter 12). The amino-terminal portion of the protein has multiple cysteine residues and a globular shape that distinguishes the glypicans from the syndecan ectodomains, which tend to be extended structures (Figure ).
A number of other membrane proteoglycans are expressed on the surface of many different cell types including the widespread CD44, NG2 (also known as CSPG4), phosphacan (PTPζ), thrombomodulin, and invariant chain of the major histocompatibility complex (MHC) class II system. Serglycin is the major cytoplasmic secretory granule proteoglycan that is present in endothelial, endocrine, and hematopoietic cells. Depending on the species, it has a variable number of glycosaminoglycan attachment sites that can carry chondroitin sulfate or heparin chains. In fact, many proteoglycans show variation in the degree of substitution by GAG chains, giving rise in some cases to so-called “part-time” proteoglycans.
To a large extent, the biological functions of proteoglycans depend on the interaction of the glycosaminoglycan chains with different protein ligands. However, the protein core determines when and where expression takes place and can itself interact with other components of the extracellular environment and the cytoskeleton. Table 38.1 lists examples of proteins known to interact with glycosaminoglycans. Proteins that bind to the sulfated glycosaminoglycan chains appear to have evolved by convergent evolution (i.e., they do not contain a specific fold present in all glycosaminoglycan-binding proteins, in contrast to other groups of glycan-binding proteins). These interactions have profound physiological effects and are discussed further in Chapter 38.
LINKAGES OF GLYCOSAMINOGLYCANS TO PROTEINS
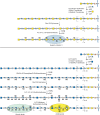
Different subtypes of sulfated glycosaminoglycans are attached to their core proteins by unique linkages. There are two types of keratan sulfate, distinguished by the nature of their linkage to protein (Figure ). KS I, originally detected in cornea, is found on an N-glycan linked to protein through an asparagine residue (Chapter 9). KS II (skeletal keratan sulfate) is found on an O-glycan core 2 structure and is thus linked through N-acetylgalactosamine to serine or threonine (Chapter 10). The structural features in control of keratan sulfate substitution remain unclear, as the underlying poly-N-acetyllactosamine backbone can be found on many other glycoproteins. Notably, in humans and bovine, the large chondroitin sulfate proteoglycan found in cartilage (aggrecan) contains a segment of four to 23 hexapeptide repeats (E-E/L-P-F-P-S) where the keratan sulfate chains are located, whereas aggrecan in rats and other rodents lacks this motif and does not contain keratan sulfate.
Keratan sulfates (KS) contain a sulfated poly-N-acetyllactosamine chain, linked to either asparagine or serine/threonine residues. Chst1 and 5 can add sulfate groups at the indicated positions. The actual order of the various sulfated and nonsulfated (more...)
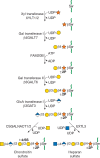
Two classes of glycosaminoglycan chains—chondroitin sulfate/dermatan sulfate and heparan sulfate/heparin—are linked to serine residues in proteins by way of xylose (Figure ). Xylosyltransferase initiates the process using UDP-xylose as donor. Two isoforms of the enzyme are known in vertebrates (XYLT1 and XYLT2), but only one isozyme exists in C. elegans and D. melanogaster. A glycine residue invariably lies to the carboxy-terminal side of the serine attachment site, but a perfect consensus sequence for xylosylation does not exist. At least two acidic amino acid residues are usually present, and they can be located on one or both sides of the serine, usually within a few residues. Several proteoglycans contain clustered glycosaminoglycan attachment sites, raising the possibility that xylosyltransferase could act in a processive manner. Xylosylation is an incomplete process in some proteoglycans, which may explain why proteoglycans with multiple potential attachment sites contain different numbers of chains in different cells.
The biosynthesis of chondroitin sulfate (left chain) and heparan sulfate (right chain) is initiated by the formation of a linkage region tetrasaccharide (green circles in Fig. 17.1). Addition of the first hexosamine commits the intermediate to either (more...)
After xylose addition, a linkage tetrasaccharide assembles by the transfer of two galactose residues catalyzed by unique members of the β4 galactosyl-, β3 galactosyl-, and β3 glucuronosyltransferase families of enzymes (Figure ). This intermediate can undergo phosphorylation at the C-2 position of xylose and in the case of chondroitin sulfate, sulfation of the galactose residues. In general, phosphorylation and sulfation occur substoichiometrically, but phosphorylation may be transient. Phosphorylation occurs early in the assembly process and creates the preferred substrate for B4GALT7; a phosphatase removes the phosphate at a later stage of biosynthesis. The function of galactose sulfation in chondroitin sulfate remains unclear.
The linkage tetrasaccharide lies at a bifurcation in the biosynthetic pathway: addition of β4-linked N-acetylgalactosamine, which initiates chondroitin sulfate assembly, or addition of α4-linked N-acetylglucosamine, which initiates heparan sulfate assembly (Figure ). Genetic evidence from studies of C. elegans suggests that N-acetylgalactosamine addition during chondroitin assembly is mediated by the same enzyme that is involved in chain polymerization (Sqv5), but biochemical evidence suggests that more than one enzyme may exist in vertebrates. In heparin/heparan sulfate formation, the addition of the first N-acetylglucosamine residue is catalyzed by an enzyme called EXTL3, which differs from the transferases involved in heparan polymerization (called EXT1 and EXT2). These enzymes are important control points because they ultimately regulate the type of glycosaminoglycan chain that will assemble. Control of the addition of β4GalNAc or α4GlcNAc appears to be manifested at the level of enzyme recognition of the polypeptide substrate.
GLYCOSAMINOGLYCAN BIOSYNTHESIS
Keratan Sulfate
Keratan sulfate chains contain a mixture of nonsulfated (Galβ4GlcNAcβ3), monosulfated (Galβ4GlcNAc6Sβ3), and disulfated (Gal6Sβ4GlcNAc6Sβ3) disaccharide units (Figure ). The biosynthesis of the poly-N-acetyllactosamine backbone is described in Chapter 14. At least two classes of sulfotransferases, one or more N-acetylglucosaminyl 6-O-sulfotransferases (e.g., CHST4 or CHST6), and one or two galactosyl 6-O-sulfotransferases (CHST1 and CHST3) catalyze the sulfation reactions. These enzymes, like other sulfotransferases, use activated sulfate (PAPS [3′-phosphoadenyl-5′-phosphosulfate]) as a high-energy donor (Chapter 5). 6-O-Sulfation of N-acetylglucosamine occurs on the nonreducing terminal residue, encouraging further chain elongation, whereas sulfation of galactose residues takes place on nonreducing terminal and internal galactose residues, with a preference for galactose units adjacent to a sulfated N-acetylglucosamine residue. Sulfation of a nonreducing terminal galactose residue blocks further elongation of the chain, providing a potential mechanism for controlling chain length. The poly-N-acetyllactosamine chains of KS I are generally longer than those of KS II and may contain up to 50 disaccharide units (20–25 kDa). The chains can be fucosylated and sialylated as well (Chapter 14).
Chondroitin Sulfate
Vertebrate chondroitin sulfate consists of repeating sulfate-substituted GalNAcβ4GlcAβ3 disaccharides polymerized into long chains (Figure ). In contrast, invertebrates such as C. elegans and D. melanogaster make either nonsulfated or low sulfated chains. The assembly process for the backbone appears to be highly conserved, based on the presence of homologous genes for all of the reactions (Chapters 25 and 26). As described above, the assembly process is initiated by the transfer of GalNAcβ3 to the linkage tetrasaccharide (Figure ). In both vertebrates and invertebrates, the polymerization step is catalyzed by one or more bifunctional enzymes (chondroitin synthases) that have both β3 glucuronosyltransferase and β4 N-acetylgalactosaminyltransferase activities. Vertebrates also express homologs that can transfer individual sugars to the chain. Chondroitin polymerization also requires the action of the chondroitin polymerizing factor (CHPF), a protein that lacks independent activity but collaborates with the polymerases to enhance the formation of polymers. Sulfation of chondroitin in vertebrates is a complex process, with multiple sulfotransferases involved in 4-O-sulfation and 6-O-sulfation of N-acetylgalactosamine residues (Figure ).
Biosynthesis of chondroitin sulfate/dermatan sulfate involves the polymerization of N-acetylgalactosamine and glucuronic acid units and a series of modification reactions including O-sulfation and epimerization of glucuronic acid to iduronic acid. Heparan (more...)
Additional enzymes exist for epimerization of D-glucuronic acid to L-iduronic acid in dermatan sulfate (DSE1-2), sulfation at the C-2 position of the uronic acids, and other patterns of sulfation found in unusual species of chondroitin (Table ). The location of sulfate groups is easily assessed using bacterial chondroitinases (ABC, B, and ACII) that cleave the chains into disaccharides. Many chains are hybrid structures containing more than one type of chondroitin disaccharide unit. For example, dermatan sulfate is defined as having one or more iduronic acid–containing disaccharide units (chondroitin sulfate B) as well as glucuronic acid–containing disaccharides (chondroitin sulfate A and C). Animal cells also degrade chondroitin sulfate in lysosomes using a series of exolytic activities (Chapter 44).
Types of chondroitin sulfates
Heparan Sulfate
Heparan sulfate assembles as a copolymer of GlcNAcα4GlcAβ4 (Figure ), which then undergoes extensive modification reactions, catalyzed by at least four families of sulfotransferases and one epimerase. N-acetylglucosamine N-deacetylase/N-sulfotransferases (NDST1-4) act on a subset of N-acetylglucosamine residues to generate N-sulfated glucosamine (GlcNSO3) units, many of which occur in clusters along the chain. Generally, the enzyme deacetylates N-acetylglucosamine and rapidly adds sulfate to the free amino group to form GlcNSO3, but a small number of glucosamine residues with unsubstituted amino groups may arise from incomplete N-sulfation. An epimerase (GLCE), different from the one involved in dermatan sulfate synthesis, then acts on some glucuronic acid residues, followed by 2-O-sulfation of some of the iduronic acid units (catalyzed by HS2ST). Some glucuronic acid units also undergo 2-O-sulfation by the same enzyme. The addition of 2-O-sulfate groups to glucuronic or iduronic acid prevents the reversible epimerization reaction. Next, 6-O-sulfotransferases (HS6ST1-3) add sulfate groups to selected glucosamine residues. Finally, certain subsequences of sulfated sugar residues and uronic acid epimers provide targets for 3-O-sulfotransferases (HS3ST1-6).
Unlike other glycosaminoglycans, heparan sulfate is further modified once displayed at the plasma membrane. A family of plasma membrane endosulfatases (SULFs) can remove sulfate groups from internal 6-O-sulfated glucosamine residues in heparan sulfate and a heparanase can cleave the chains at limited sites. This post-assembly processing of the chains at the cell surface results in altered response of cells to growth factors and morphogens. Intriguingly, the mammalian genome contains other sulfatases of unknown function, raising the possibility that other post-assembly processing reactions of glycosaminoglycans may occur.
In contrast to chondroitin chains, which tend to have long tracts of fully modified disaccharides, the modification reactions in heparan sulfate biosynthesis occur in clusters along the chain, with regions devoid of sulfate separating the modified domains. In general, the reactions proceed in the order indicated, with evidence for functional interdependence within and between the biosynthetic enzyme families, but they often fail to go to completion, resulting in tremendous chemical heterogeneity.
Readout and Regulation of Glycosaminoglycan Assembly
The disaccharide composition of glycosaminoglycan chains can be readily assessed using bacterial lyases or chemical degradation methods (which are more useful for differentiating glucuronic and iduronic acids). Direct sequencing of the chains has proved difficult because of their heterogeneity, however the application of specific lysosomal exoenzymes involved in glycosaminoglycan degradation and new mass spectrometry methods are making significant inroads into sequencing of glycosaminoglycans (Chapter 50). The specific arrangement of sulfated residues and uronic acid epimers in heparin/heparan sulfate and dermatan sulfate gives rise to binding sequences for proteins. The three examples shown in Figure show sequences that can interact with antithrombin, heparin cofactor II, and other potential proteins. More modified sequences can interact as well, and the binding of many ligands is dependent on the presence of correctly orientated charged clusters rather than the specific presence of individual sulfated groups. Binding of glycosaminoglycans to proteins is described in greater detail in Chapter 38. A major question remains regarding how the enzymes and biosynthetic pathways are regulated to achieve tissue-specific expression of protein-binding sequences.
During the last decade, most if not all of the enzymes involved in glycosaminoglycan synthesis have been purified and molecularly cloned from mammals and model organisms. Several important features have emerged from these studies, which may shed light on how different protein-binding sequences arise.
Several of the enzymes appear to have dual catalytic activities. Thus, a single protein bearing two catalytic domains catalyzes N-deacetylation of N-acetylglucosamine residues and subsequent N-sulfation (NDSTs) in heparan sulfate formation. The same is true of the copolymerases, which transfer N-acetylglucosamine and glucuronic acid (heparan sulfate) and N-acetylgalactosamine and glucuronic acid (chondroitin sulfate) from the corresponding UDP sugars to the growing polymer. In contrast, the epimerases and O-sulfotransferase activities appear to be unique properties of independent enzymes.
In several cases, multiple isozymes exist that can catalyze either a single or a pair of reactions. Thus, four N-deacetylase/N-sulfotransferases, three 6-O-sulfotransferases, seven 3-O-sulfotransferases and two endo-sulfatases (SULFs) have been identified in heparan sulfate biosynthesis. Their tissue distribution varies and differences exist in substrate preference, which may cause differences in the pattern of sulfation. However, some overlap in expression and in substrate utilization occurs as well. Multiple isozymes of 4-O- and 6-O-sulfotransferases also can participate in chondroitin sulfate formation.
The polymerization and polymer modification reactions probably colocalize in the same stacks of the Golgi complex. Thus, the enzymes may form supramolecular complexes that coordinate these reactions. The composition of these complexes may play a part in regulating the fine structure of the chains.
In general, the composition of heparan sulfate, and likely chondroitin and dermatan sulfate, on a given proteoglycan varies more between cell types than on different core proteins expressed in the same cell. This observation suggests that each cell type may express a unique array of enzymes and potential regulatory factors. The mechanisms behind the generation of apparently cell-specific glycosaminoglycan chains through regulated, yet partly stochastic modification reactions are yet to be fully elucidated. However, recently identified transcriptional regulation of key heparan sulfate biosynthetic enzymes suggests we may be close to this understanding.
Recombinant enzymes and new synthetic schemes are increasingly used to generate defined glycosaminoglycan oligosaccharides, which can be used to probe ligand-binding affinities and specificities. Recent innovations include re-engineering biosynthetic enzymes to introduce tailored specificities, allowing the creation of glycosylation patterns not seen in nature and the exploration of their biological impact.
Arrays of synthetic glycosaminoglycans used to probe the importance of chain length, sulfation pattern, and chain density are now in common use and enable the rapid screening of glycosaminoglycan–protein interactions. As highlighted above, care needs to be taken when inferring a biological effect from these interactions. However, recently developed chemical strategies for creating artificial proteoglycans at the cell surface together with new opportunities for altering cell surface display of glycosaminoglycans using gene editing, or by application of soluble inhibitors/activators to influence components of the biosynthetic pathway, offer controlled environments with which to test glycosaminoglycan–protein interactions at the cellular level.
HEPARIN VERSUS HEPARAN SULFATE
Considerable confusion exists regarding the definition of heparin and heparan sulfate (even the spelling is often in error!). Heparin is produced by a limited number of cells, notably connective tissue-type mast cells and bipotential glial progenitor cells, whereas heparan sulfate is made by virtually all types of cells. During biosynthesis, heparin undergoes more extensive sulfation and uronic acid epimerization, such that >80% of the N-acetylglucosamine residues are N-deacetylated and N-sulfated and >70% of the glucuronic acids undergo epimerization to iduronic acid. Heparin derived from porcine entrails and bovine lung is prepared commercially by selective precipitation and is sold by pharmaceutical companies as an anticoagulant because of its high capacity to bind to antithrombin (Chapters 38 and 57). The active sequence is a pentasaccharide shown in Figure , which is now sold as a purely synthetic anticoagulant (Arixtra). Low-molecular-weight heparins are derived from commercial unfractionated heparin by chemical or enzymatic cleavage, depending on the brand. Selectively desulfated forms of heparin and heparin oligosaccharides are also available commercially, some of which lack anticoagulant activity, but still retain other potentially useful properties. Heparan sulfate also can contain anticoagulant activity, but typical preparations from cells or tissues are much less active than heparin. Care should be taken in extrapolating data obtained with heparin (e.g., binding of a protein to heparin-Sepharose) versus binding to heparan sulfate and heparan sulfate proteoglycans; binding to heparin can occur owing to the high charge content of the polysaccharide, whereas the same factor might bind to heparan sulfate with lower affinity or not at all. On the other hand, specific protein-binding motifs expressed in subspecies of heparan sulfate may occur also in heparin, although concealed by additional, redundant, sulfate residues.
PROTEOGLYCAN PROCESSING AND TURNOVER
Cells secrete matrix proteoglycans directly into the extracellular environment (e.g., members of the aggrecan family, the basement membrane proteoglycans, SLRPs, and serglycin). However, others are shed from the cell surface through proteolytic cleavage of the core protein through matrix metalloproteases (e.g., the syndecans).
Extracellular heparanase, an endo-β-glucuronidase, cleaves heparan sulfate at restricted sites resulting in release of growth factors or chemokines immobilized on heparan sulfate proteoglycans at cell surfaces or in the ECM. The activity of shedding extracellular domains can contrast with that of the intact transmembrane proteoglycan and digestion by heparanase can similarly generate glycosaminoglycan fragments with distinct and contrasting activity. This is a particular feature of invading cells in which secreted heparanase can act in concert with matrix metalloproteases to remodel the ECM. Because heparanase has been implicated in the progression of various cancers, there is considerable interest in the development of inhibitors. This has led to clinical trials involving inhibitors either alone or in combination with other therapeutic agents (Chapter 47).
Cells also internalize a large fraction of cell-surface heparan sulfate proteoglycans by endocytosis. These internalized proteoglycans first encounter proteases and heparanase and the core protein and the heparan sulfate chains are cleaved endolytically. The resulting smaller heparan sulfate fragments eventually appear in the lysosome and undergo complete degradation by way of a series of exoglycosidases and sulfatases (Chapter 44). Chondroitin sulfate and dermatan sulfate proteoglycans follow a similar endocytic route. One of the human hyaluronidases (HYAL-4) has been found to be involved in endolytic degradation of chondroitin sulfate.
ACKNOWLEDGMENTS
The authors appreciate helpful comments and suggestions from Barbara Mulloy.
FURTHER READING
Mizumoto S, Yamada S, Sugahara K. 2015. Molecular interactions between chondroitin-dermatan sulfate and growth factors/receptors/matrix proteins. Curr Opin Struct Biol
34: 35–42. doi:10.1016/j.sbi.2015.06.004 [
PubMed: 26164146] [
CrossRef]
Mitsou I, Multhaupt HM, Couchman JR. 2017. Proteoglycans, ion channels and cell-matrix adhesion. Biochem J
474: 1965–1979. doi:10.1042/BCJ20160747 [
PubMed: 28546458] [
CrossRef]
Kjellén L, Lindahl U. 2018. Specificity of glycosaminoglycan–protein interactions. Curr Opin Struc Biol
50: 101−108. doi:10.1016/j.sbi.2017.12.011 [
PubMed: 29455055] [
CrossRef]
Annaval T, Wild R, Crétinon Y, Sadir R, Vivès RR, Lortat-Jacob H. 2020. Heparan sulfate proteoglycans biosynthesis and post synthesis mechanisms combine few enzymes and few core proteins to generate extensive structural and functional diversity. Molecules
25: 4215. doi:10.3390/molecules25184215 [
PMC free article: PMC7570499] [
PubMed: 32937952] [
CrossRef]