

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Probe Reports from the NIH Molecular Libraries Program [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2010-.

Cryptococcus neoformans is pathogenic yeast that primarily causes pulmonary and central nervous system infections in immune-compromised patients. It is most recognized as a cause of morbidity and mortality in HIV patients and kills approximately 600,000 people annually with the majority of deaths in resource-limited regions with high HIV endemicity. One of the major medical issues for the treatment of C. neoformans is that the gold standard therapy of amphotericin B plus 5-flucytosine is not available in resource-limited regions where it is most desperately needed. The success of this therapy is due to its ability to directly kill the fungus in the CNS (fungicidal activity). The therapy most widely used in resource-limited regions is fluconazole, which is a fungistatic agent which, although it does directly kill Cryptococcus to a limited extent, is associated with poorer outcomes as well the emergence of drug-resistant organisms. Consequently, new agents are needed for the treatment of this pathogen.
Only one new class of antifungal agents has been developed in the last twenty years (i.e. echinocandins) and it does not have clinically useful activity against C. neoformans. The echinocandins inhibit the synthesis of 1,3-β-glucan, the key structural component of fungus cell walls. Since the cell wall is unique to fungi with no analog in humans, it is a very attractive antifungal drug target. Despite the fact that C. neoformans synthesizes 1,3-β-glucan, the organism is resistant to echinocandins and, to date, there are very few molecules with any activity toward cell wall processes in C. neoformans. Therefore, the identification of this new molecule that targets the C. neoformans cell wall not only provides a potential antifungal drug candidate but is also a useful mechanistic probe with which to study the unique cell wall biosynthesis machinery in this pathogen.
Assigned Assay Grant #: R01 AI091422
Screening Center Name & PI: Broad Institute Probe Development Center, Stuart Schreiber, PhD
Chemistry Center Name & PI: Broad Institute Probe Development Center, Stuart Schreiber, PhD
Assay Submitter & Institution: Damian Krysan, PhD, University of Rochester, NY
PubChem Summary Bioassay Identifier (AID): 651665
Probe Structure & Characteristics

ML407
| CID/ML# | Target Name | IC50/EC50 [SID, AID] | Anti-target | IC50/EC50 [SID, AID] | Fold Selective | Secondary Assays: IC50/EC50 (nM) [SID, AID] |
|---|---|---|---|---|---|---|
| CID 970188/ML407 | C. neoformans cell wall | 595 nM* [SID 163621169; AID 687000, AID 743159, AID 743303, AID 743302] | Sorbitol-protected/sorbitol-unprotected | 9.9 μM/5.6 μM* [SID 163621169; AID 686997, AID 686999, AID 743123, AID 743298, AID 743128, AID 743305, AID 743304] | 1.8 | Cytotoxicity HepG2 inactive; A549 inactive [SID 163621169; AID 687003, AID 687002, AID 743315, AID 743131, AID 743316, AID 743129, AID 743311, AID 743313, AID 743158, AID 743312, AID 743318] |
- *
represents an average across several batches
1. Recommendations for Scientific Use of the Probe
This probe development project was designed to identify small molecules that interfere with cell wall processes in the pathogenic yeast Cryptococcus neoformans. The secondary screening assays indicate whether the molecular scaffold identified by this effort interferes with signaling through the cell wall integrity MAP kinase pathway. Although the proteins of this pathway are conserved across most fungi, recent studies indicate that there are features of the pathway in Cryptococcus neoformans that are unique. The initial kinase in the cell wall pathway is Pkc1, a homologue of the well-known mammalian kinase, protein kinase C (1). A variety of stressors, including growth at elevated temperature, oxidative/nitrosative conditions and direct cell wall damage, activate Pkc1 to initiate signaling through the MAP kinase cascade (2). The ultimate output of this pathway is the expression of a large set of genes involved in cell wall integrity.
This pathway has been studied frequently using the promiscuous protein kinase inhibitor staurosporine that inhibits Pkc1 in all forms of yeast (1). However, this kinase inhibitor has activity against a large set of kinases and is very toxic to host cells, limiting its utility in the study of this pathway in the setting of infection. Other Pkc1 inhibitors have been reported but have similar drawbacks and most are not readily available (3). Recently, the Lodge group has also shown that, in contrast to all other fungi studied to date, the cell wall integrity pathway in C. neoformans is activated by heat stress in a Pkc1-independent fashion (4), suggesting that the inhibitor molecules currently most widely used to study cell wall integrity pathway signaling (Pkc1) will not interrogate some important functions of the pathway.
The probe identified in this project will allow investigators to interrogate the role of cell wall integrity signaling in C. neoformans and, possibly, other fungi in a wide variety of settings. Thus, it should have utility to investigators interested in studying fungal cell wall integrity. Based on previous uses of molecules like staurosporine, it should be a more specific way to: 1) identify and study mutants with defects in cell wall biosynthesis and signaling; 2) perform chemical genetic analyses to identify pathways that interact with the cell wall integrity pathway; 3) interrogate cell wall stress responses and their effects in vivo; and 4) one could use the probe as a means to increase the sensitivity of high throughput chemical screens for molecules that interfere with cell wall biosynthesis.
One of the most exciting prospects for this class of probe is related to its lack of toxicity toward mammalian cells. This opens up the possibility of using the probe to block cell wall stress responses during interactions with host cells. In particular, C. neoformans is able to replicate within the phagolysosome of macrophages; however, it has been difficult to study this process since strains with mutations in cell wall genes have altered phagocytosis or are rapidly killed. Using a non-toxic inhibitor of cell wall stress responses, would allow one to modulate cell wall signaling pathway activity in the presence of host cells.
2. Materials and Methods
Cell Lines
The following cell lines were used in this study:
- Cryptococcus neoformans var. grubii H99 (ATCC #208821); wild-type strain used for primary assay
- Saccharomyces cerevisiae (Krysan lab) containing plasmid with Rlm1 gene; used for Rlm1 and Calcofluor White assays
- HEK293 mammalian kidney cells (ATCC; CRL-1573); used for mammalian toxicity assays
- HEPG2 mammalian liver cells (ATCC; 77400); used for mammalian toxicity assays
- HeLa cervical mammalian cells (ATCC; CCL-2); used for mammalian toxicity assays
- A549 human lung cancer cells (ATCC; CCL-185); used for mammalian toxicity assays
2.1. Assays
2.1.1. Primary C. neoformans HTS (AID 651654) and C. neoformans Dose-Response Retest (AID 687000)
The primary assay measures adenylate kinase levels in the culture media (as measured by increased luminescence in the presence of ToxiLight reagent) by potential cell wall damaging compounds. The assay was performed using a wild-type Cryptococcus inoculums, dispensed into 384-well assay plates (1536-well for the primary assay) into which 100 nL of compound have been pinned (7.5 nL for the primary assay). After 4 hours of incubation at 37 degrees C, ToxiLight reagent is added. The plates are shaken for 5 minutes, then held for 30 minutes before luminescence output is read. Compounds possessing an IC50 value less than 10 μM were selected for further studies.
2.1.2. Counter screen Mammalian Cell Toxicity Assays (AID 687005, AID 720491, AID 687003, AID 687002, AID 687004)
A mammalian cell panel was subjected to incubation with compounds of interest for 72 hours. Subsequent cell viability was determined via measuring luminescence using Cell Titer Glo reagent. Toxicity at less than 20 μM was considered to be below the acceptable threshold and resulted in exclusion of the associated compound from further consideration.
2.1.3. Secondary Antifungal IC50 with Alamar Blue (AID 686999) and Counter screen for detection of yeast cell lysis in presence of Sorbitol (AID 686997)
Sorbitol confers osmotic protection to the Cryptococcus cell wall, presenting a differential between IC50 values between cells grown in sorbitol and those without when there is damage to the cell wall. Cells are added to 384-well plates and compound is pinned into the wells. Plates are incubated at 37C for 24 hours. Alamar blue is added to all wells and the plates are further incubated for 3 hours before reading for fluorescent output.
2.1.4. Secondary Rlm1 Stress Factor Reporter Assay in S. cerevisiae (AID 743130) and Counterscreen Calcofluor White Reporter Assay in S. cerevisiae (AID 743124)
These are the same assays, run in the presence or absence of Calcofluor White. The Rlm1 stress factor assay uses a cell wall stress reporter strain of S. cerevisiae which contains a plasmid encoding multiple copies of the Rlm1 consensus binding sites fused to beta-galactosidase. Rlm1 is regulated by the cell wall integrity protein kinase pathway that is conserved in C. neoformans and cells subject to cell wall stress transcribe this gene. An increase of the readout signal indicates an up-regulation of the cell wall stress response pathway. Results are compared to those in the Calcofluor White assay. A decrease of signal in the Calofluor White assay indicates that the cell wall stress response is diminished and less Rlm1 is being produced, probably due to signaling pathway interference. Calcofluor White is a less-specific way to look at what is happening with the cell wall. For these assays, log phase S. cerevisiae is incubated with compounds of interest in the presence or absence of Calcofluor White for 4 hours at room temperature. Gal-screen is added, and after another hour at room temperature the plates are read for luminescence output.
2.1.5. Secondary Phosphorylation of C. neoformans Mpk1 (AID 743392)
Cell wall stress in C. neoformans leads to the phosphorylation of the MAP kinase Mpk1. This assay is used as a confirmation of cell wall stress activation in the S. cerevisiae-based Rlm1/Calcofluor White assays by assessing the phosphorylation of Mpk1 during cell wall stress due to Calcofluor White, using Western blots. Relative densities of the bands are determined and percentage decrease of phosphorylation is visualized with the ECL Plus Western blotting detection system.
2.2. Probe Chemical Characterization
Table 1Summary of Known Probe Properties in PubChem
| IUPAC Chemical Name | 3-[2-(5-methyl-2-propan-2-ylphenoxy)ethyl]-1H-benzimidazole-2-thione |
|---|---|
| PubChem CID | CID 970188 |
| Molecular Weight | 326.45578 [g/mol] |
| Molecular Formula | C19H22N2OS |
| XLogP3-AA | 4.5 |
| H-Bond Donor | 1 |
| H-Bond Acceptor | 2 |
| Rotatable Bond Count | 5 |
| Exact Mass | 326.145284 |
| Topological Polar Surface Area | 56.6 |
ML407 was prepared as described in Section 2.3, and was analyzed by UPLC, 1H and 13C NMR spectroscopy, and high-resolution mass spectrometry. The data obtained from NMR and mass spectrometry were consistent with the structure of the probe, and UPLC analysis showed purity of >90%.
The solubility of the probe was determined to be 0.18 μM in phosphate-buffered saline (PBS; pH 7.4, 23°C) solution with 1% DMSO. The probe is stable in PBS (1% DMSO), with 97.6 % remaining after 48 hours (Figure 1). The compound is also very stable in human and mouse plasma, with 100% remaining after a 5-hour incubation period. Plasma protein binding (PPB) studies showed that it was 100% bound in human plasma, and 100% bound in mouse plasma.
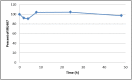
Figure 1
Stability of Probe (ML407) in PBS Buffer (with 1% DMSO, pH 7.4, 23°C)*.
2.3. Probe Preparation
The probe (ML407) and most of the analogs prepared for structure-activity relationship analysis were prepared by the synthetic sequence outlined below in Figure 2. 2-Isopropyl-5-methylphenol 1 was alkylated with tert-butyl (2-bromoethyl)carbamate 2 to provide intermediate 3, which was deprotected with trifluoroacetic acid (TFA) and the resulting free amine was then reacted with 1-fluoro-2-nitrobenzene 4 to afford 5. The nitro group was reduced by hydrogenation to afford aniline 6, which was cyclized with di(1H-imidazol-1-yl)methanethione to provide the probe 7 (ML 407).
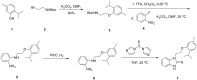
Figure 2
Synthesis of probe ML407.
3. Results
ML407 displays dose dependent cell wall damage in wild type wild-type Cryptococcus neoformans as measured by adenylate kinase levels in the culture media (Figure 3A; IC50 = 595 nM) and displays no cytotoxicity to A5449 cells (Figure 3B). Addition of sorbitol provides some protection to cell damage caused by ML407 (Figure 4A and 4B). The cell wall stress induced by ML407 was also confirmed by Rlm1 response and Calcofluor White cell signaling response (Figure 5).
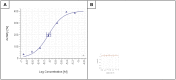
Figure 3
C. neoformans cell wall damage (AID 743159), AbsAC40 = 0.251 μM (A); A549 cytotoxicity (AID 743158 ), AC50 = >12 μM (B).
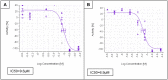
Figure 4
C. neoformans sorbitol plus assay (AID 743298), AC50 = 9.6 μM (A); C. neoformans sorbitol minus assay (AID 743304, AID 743305), AC50 = 6.9μ(B).

Figure 5
S. cerevisiae Rlm1 response assay (AID 743130), absAC40 = 27 μM (A) and S. cerevisiae Calcofluor White assay (AID 743124), AC50 = 35 μM (B).
3.1. Dose Response Curves for Probe
Figure 3 Dose-dependent Activity of the probe (ML407) in the AK release assay (A) with no toxicity in mammalian A549 cells (B)
Figure 4 Sorbitol protective effect on C. neoformans cell wall in the presence of ML407 (A) as compared to inhibition without sorbitol (B).
Figure 5 S. cerevisiae Rlm1 response (AID 743130) (A) and S. cerevisiae Calcofluor White cell signaling response (AID 743124) (B)
3.2. Cellular Activity
All the assays performed in this project are cell based assays
3.3. Profiling Assays
None
4. Discussion
4.1. Comparison to Existing Art and How the New Probe is an Improvement
We compared the performance of ML407 with Staurosporine and OSU-03012. These results are presented in Table 2. ML407 is superior to Staurosporine and OSU-03012. Staurosporine is a non-specific protein kinase inhibitor and it is too toxic to function in the primary assay and did not show selectivity in the sorbitol +/- assay. OSU-03012 was originally developed as anti-cancer agent and the assay provider later discovered its anti-cryptococcal activity through inhibition of Phosphoinositide-dependent kinase-1 (PDK1) orthologues. (5) It is also cytotoxic and is 10 fold less potent than ML407. ML407 demonstrated submicromolar activity in the primary assay and showed approximately 2-fold selectivity in the sorbitol +/- assay. This probe compound is non-cytotoxic towards mammalian cells. ML407 is a cell wall stress signaling inhibitor and has target-related biological mode of action.
Table 2
Comparison of Probe ML407 to selected Prior Art Compounds.
This initial hit compound (CID 773463) has been tested in 496 PubChem assays. Of those, it was active in 24 assays, 6 of which were for the present C. neoformans screening project.
There are a few compounds reported in the literature with cell wall targeted anti-cryptococcal activity in C. neoformans. Fungal chitin synthase inhibitors such as pentoxifylline (10) and 2′-benzoyloxycinnamaldehyde (11) could rupture the cell wall with an IC50 of 10 μM and a MIC of 16 μg/mL, respectively. The natural product, nikkomycin Z (12) can also inhibit chitin synthesis. Caspofungin, a β-1,3-glucan synthase inhibitor, is relatively ineffective against C. neoformans (12). The assay provider used Caspofungin as positive control in their initial assay development (Figure 6).
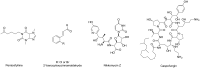
Figure 6
Examples of compounds with cell wall targeted anti-cryptococcal activity.
The assay provider reported that staurosporine, the canonical nonspecific protein kinase inhibitor, is highly toxic to both mammalian and fungal cell. The MIC of straurosporine against C. neoformans is about 0.6 μg /mL. The assay provider also suggested the use of colistin (polymixin), a less expensive antibiotic, as positive control. The assay provider discovered that Tamoxifen and Clomiphene caused cell lysis in C. neoformans with a MIC of 64 μg /mL (13) (Figure 7). Tamoxifen was used as positive control in the pilot screening and 8 compounds with in vitro activity at concentrations < 64 μg /mL were identified in the screen. The structures of those compounds have not been released. The assay provider also reported three Phosphoinositide-Depdendent -1 Kinase (PKI) inhibitiors, KP-372-1, OSU-03012 and UCN-01, showed anti-cryptococcal activity (5). However, these molecules have significant limitations as specific mechanistic probes. KP-372-1 inhibits at least three classes of kinases, including PDK1 and protein kinase B; since there are homologues of each in yeast, KP-372-1 is not a useful mechanistic probe (6). OSU-03012 also appears to inhibit PDK1 orthologues but has been shown to have activity against p21-activated protein kinases (7), which are essential in C. neoformans. Accordingly, the mechanistic utility of this molecule is reduced by its lack of specificity; in addition, the assay provider has new data indicating that OSU-03012 analogs interfere with vacuolar fusion and biogenesis. In our hands, we have found it to be broadly cytotoxic.
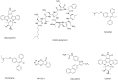
Figure 7
Compounds identified with cell wall targeted anti-cryptococcal activity in the pilot screen.
In conclusion, there are few potent chemical entities with cell wall targeted anti-cryptococcal activity in C. neoformans. Thus, research to find new scaffolds with cell wall damaging anti-cryptococcal activity is warranted.
5. References
- 1.
- Levin D. Cell wall integrity signaling in S. cerevisiae. Microbiol. Mol. Biol. Rev. 2005;69:262–291. [PMC free article: PMC1197416] [PubMed: 15944456]
- 2.
- Gerik KJ, Donlin MJ, Soto CE, Banks AM, Maligie MA, Selitrennikoff CP, Lodge JK. Cell wall integrity is dependent on the Pck1 signal transduction pathway in Cryptococcus neoformans. Mol. Microbiol. 2005;58:393–408. [PubMed: 16194228]
- 3.
- Sussman A, et al. Discovery of cercosporamide, a known antifungal natural product, as a selective Pkc1 inhibitor through high throughput screening. Eukaryot Cell. 2004;3:932–943. [PMC free article: PMC500880] [PubMed: 15302826]
- 4.
- Lam WC, Gerik KJ, Lodge JK. Role of Cryptococcus neoformans Rho1 GTPases in the PKC1 signaling pathway in response to thermal stress. Eukaryot Cell. 2013;12:118–131. [PMC free article: PMC3535842] [PubMed: 23159519]
- 5.
- Baxter BK, DiDone L, Oga D, Schor S, Krysan DJ. Identification, in vitro activity and mode of action of phosphoinositide-1 kinase inhibitors as antifungal molecules. ACS Chem Biol. 2011;6:502–510. [PMC free article: PMC3098953] [PubMed: 21294551]
- 6.
- Zeng Z, et al. Simultaneous inhibition of PDK1/Akt and Fms-like tyrosine kinase 3 signaling by a small-molecule KP372-1 induces mitochondrial dysfunction and apoptosis in acute myelogenous leukemia. Cancer Res. 2006;66:3737–3746. [PubMed: 16585200]
- 7.
- May Y, et al. Development of p21-activated kinase targeted multikinase inhibitors that inhibit thyroid cancer cell migration. J Clin Endocrinol Metab. 2013 [PMC free article: PMC3733855] [PubMed: 23709653] [CrossRef]
- 8.
- Krysan DJ, DiDone L. A high-throughput screening assay for small molecules that disrupt yeast cell integrity. J Biomol Screen. 2008;181:1073–81. [PubMed: 18626115]
- 9.
- Harrison D, Ralph JT. The infrared spectra of some benzimidazoline-2-thiones and benzimidazol-2-yl sulphides. J.Chem.Soc. (B). 1967:14–15.
- 10.
- Tsirilakis K, et al. Methylxanthine inhibit fungal chitinases and exhibit antifungal activity. Mycopathologia. 2012;173:83–91. [PMC free article: PMC4289597] [PubMed: 21968902]
- 11.
- Tae hoon K, et al. Inhibition of chitin synthases and antifungal activities by 2′- benzoyloxyloxycinnamaldehyde from Pleuropterus ciliinervis and its derivatives. Biol. Pharm. Bull. 2007;30(3):598–602. [PubMed: 17329866]
- 12.
- Peter RK, et al. The Cryptococcus neoformans MAP kinase Mpk1 regulates cell integrity in response to antifungal drugs and loss of calcineurin function. Molecular Microbiology. 2003;48(5):1377–1387. [PMC free article: PMC1635492] [PubMed: 12787363]
- 13.
- Kristy D, et al. Antifungal activity of Tamoxifen: In vitro and In vivo activities and mechanistic characterization Antimicrobial agents and chemotherapy. Antimicrob Agents Chemother. 2009:3337–3346. [PMC free article: PMC2715577] [PubMed: 19487443]
- PMCPubMed Central citations
- PubChem SubstanceRelated PubChem Substances
- PubMedLinks to PubMed
- Review In vitro antifungal activities of anidulafungin and micafungin, licensed agents and the investigational triazole posaconazole as determined by NCCLS methods for 12,052 fungal isolates: review of the literature.[Rev Iberoam Micol. 2003]Review In vitro antifungal activities of anidulafungin and micafungin, licensed agents and the investigational triazole posaconazole as determined by NCCLS methods for 12,052 fungal isolates: review of the literature.Espinel-Ingroff A. Rev Iberoam Micol. 2003 Dec; 20(4):121-36.
- Astemizole and an analogue promote fungicidal activity of fluconazole against Cryptococcus neoformans var. grubii and Cryptococcus gattii.[Med Mycol. 2010]Astemizole and an analogue promote fungicidal activity of fluconazole against Cryptococcus neoformans var. grubii and Cryptococcus gattii.Vu K, Gelli A. Med Mycol. 2010 Mar; 48(2):255-62.
- Review Toward improved anti-cryptococcal drugs: Novel molecules and repurposed drugs.[Fungal Genet Biol. 2015]Review Toward improved anti-cryptococcal drugs: Novel molecules and repurposed drugs.Krysan DJ. Fungal Genet Biol. 2015 May; 78:93-8. Epub 2014 Dec 13.
- Identification of Multiple Cryptococcal Fungicidal Drug Targets by Combined Gene Dosing and Drug Affinity Responsive Target Stability Screening.[mBio. 2016]Identification of Multiple Cryptococcal Fungicidal Drug Targets by Combined Gene Dosing and Drug Affinity Responsive Target Stability Screening.Park YD, Sun W, Salas A, Antia A, Carvajal C, Wang A, Xu X, Meng Z, Zhou M, Tawa GJ, et al. mBio. 2016 Aug 2; 7(4). Epub 2016 Aug 2.
- Practice guidelines for the management of cryptococcal disease. Infectious Diseases Society of America.[Clin Infect Dis. 2000]Practice guidelines for the management of cryptococcal disease. Infectious Diseases Society of America.Saag MS, Graybill RJ, Larsen RA, Pappas PG, Perfect JR, Powderly WG, Sobel JD, Dismukes WE. Clin Infect Dis. 2000 Apr; 30(4):710-8. Epub 2000 Apr 20.
- Discovery and Evaluation of Fungicidal Anti-Cryptococcal Molecules - Probe Repor...Discovery and Evaluation of Fungicidal Anti-Cryptococcal Molecules - Probe Reports from the NIH Molecular Libraries Program
Your browsing activity is empty.
Activity recording is turned off.
See more...