

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
National Research Council (US) Subcommittee on the Tenth Edition of the Recommended Dietary Allowances. Recommended Dietary Allowances: 10th Edition. Washington (DC): National Academies Press (US); 1989.

Recommended Dietary Allowances: 10th Edition.
Show detailsBoth animal and plant proteins are made up of about 20 common amino acids. The proportion of these amino acids varies as a characteristic of a given protein, but all food proteins—with the exception of gelatin—contain some of each. Amino nitrogen accounts for approximately 16% of the weight of proteins. Amino acids are required for the synthesis of body protein and other important nitrogen-containing compounds, such as creatine, peptide hormones, and some neurotransmitters. Although allowances are expressed as protein, a the biological requirement is for amino acids.
Proteins and other nitrogenous compounds are being degraded and resynthesized continuously. Several times more protein is turned over daily within the body than is ordinarily consumed, indicating that reutilization of amino acids is a major feature of the economy of protein metabolism. This process of recapture is not completely efficient, and some amino acids are lost by oxidative catabolism. Metabolic products of amino acids (urea, creatinine, uric acid, and other nitrogenous products) are excreted in the urine; nitrogen is also lost in feces, sweat, and other body secretions and in sloughed skin, hair, and nails. A continuous supply of dietary amino acids is required to replace these losses, even after growth has ceased.
Amino acids consumed in excess of the amounts needed for the synthesis of nitrogenous tissue constituents are not stored but are degraded; the nitrogen is excreted as urea, and the keto acids left after removal of the amino groups are either utilized directly as sources of energy or are converted to carbohydrate or fat.
Nine amino acids—histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, and valine—are not synthesized by mammals and are therefore dietarily essential or indispensable nutrients. These are commonly called the essential amino acids. Histidine is an essential amino acid for infants, but was not demonstrated to be required by adults until recently (Cho et al., 1984; Kopple and Swendseid, 1981). Under special circumstances (e.g., in premature infants or in people with liver damage), amino acids such as cystine and tyrosine, not normally essential, may become so because of impaired conversion from their precursors (Horowitz et al., 1981). Arginine is synthesized by mammals but not in amounts sufficient to meet the needs of the young of most species. Although it is not believed to be required by the human infant for normal growth, the need for arginine by the premature infant is unknown. When arginine is present in small amounts relative to other amino acids (such as in intravenous solutions or amino acid mixtures), or when liver function is compromised, arginine synthesis may be insufficient for adequate function of the urea cycle (Heird et al., 1972).
GENERAL SIGNS OF DEFICIENCY
Protein deficiency rarely occurs as an isolated condition. It usually accompanies a deficiency of dietary energy and other nutrients resulting from insufficient food intake. The symptoms are most commonly seen in deprived children in poor countries. Where protein intake is exceptionally low, there are physical signs—stunting, poor musculature, edema, thin and fragile hair, skin lesions—and biochemical changes that include low serum albumin and hormonal imbalances. Edema and loss of muscle mass and hair are the prominent signs in adults. Deficiency of this severity is very rare in the United States, except as a consequence of pathologic conditions and poor medical management of the acutely ill.
GENERAL PRINCIPLES FOR ESTIMATING PROTEIN REQUIREMENTS
At submaintenance levels of protein intake, a diminished turnover of tissue protein is accompanied by a reduced catabolic rate for the amino acids liberated by protein breakdown (Young and Scrimshaw, 1977). Similarly, turnover rate is increased with increased intake. In this way, the tissue protein pool can, within limits, enter a new steady state appropriate for the diminished or increased protein intake from food.
Under the experimental conditions of a protein-free diet, protein synthesis and breakdown continue by reutilizing amino acids. This process becomes very efficient, but some amino acids are still catabolized and the nitrogen is excreted. This lower limit, termed the obligatory nitrogen loss, has been extensively studied in adults fed protein-free diets. Values are remarkably uniform. In a series of 11 studies involving more than 200 adults ranging in age from 20 to 77 years, daily obligatory nitrogen losses averaged 53 mg (41–69 mg, range of study means) of nitrogen per kilogram daily (WHO, 1985). b
In the past, a factorial method was used as a basis for predicting the protein requirements of various age groups. For adults, the requirement for dietary protein was considered to be the amount needed to replace the obligatory nitrogen loss after adjustment for inefficiency in utilization of dietary protein and for the quality of the dietary protein consumed (i.e., its digestibility and amino acid composition). For children and pregnant and lactating women, an additional amount of protein for tissue growth or milk formation was incorporated into this factorial estimate of requirements. Because of the assumptions required, the validity of the factorial approach has been questioned.
WHO (1985) reviewed the evidence on protein requirements and concluded that adult protein allowances should be based on nitrogen balance studies. For older infants and children, data are sparse; thus, all lines of evidence were used by the subcommittee in estimating requirements—nitrogen balance and observed and theoretical needs for adequate growth. For pregnancy, nitrogen balance data were considered, but allowances continue to rely on theoretical deposition of protein in the fetus and adnexa. New information on human milk volume was used to estimate lactational requirements.
Protein synthesis and breakdown are energy-dependent and thus are sensitive to dietary energy deprivation. Consequently, the body's energy balance becomes an important factor in determining nitrogen balance and influences the apparent utilization of dietary protein. Protein requirements are determined and allowances established for conditions of adequate energy intake and balance.
Nitrogen Balance
Nitrogen balance is the difference between nitrogen intake and the amount excreted in urine, feces, and sweat, together with minor losses occurring by other routes. To estimate the protein requirement, levels of dietary protein below and near predicted adequate intake are fed and nitrogen balance is measured at each level. The requirement is estimated by interpolating or extrapolating the nitrogen balance data to the zero balance point (nitrogen equilibrium) for adults or to a defined level of positive balance (to allow for growth) for children.
Because of methodologic problems, it is difficult to attain exact measurement of intake and output of nitrogen (Hegsted, 1976), to determine the time required for adjustment at altered levels of protein intake (Rand et al., 1981), and to measure or otherwise account for nitrogen losses through routes other than urine and feces.
OTHER CRITERIA OF ADEQUACY
Most studies of protein requirements have been short. Because of the methodologic problems cited above, longer studies should provide a better basis for determining protein requirements c ; they would permit the measurement of variables such as alterations in lean body mass or in growth rate of children, which respond more slowly to dietary inadequacy. In the few long-term studies that have been reported, investigators have explored the usefulness of various biochemical indices (e.g., serum aspartate and alanine amino transferase activities) (Garza et al., 1977), but no agreement on a sensitive and reliable marker has been reached (Solomons and Allen, 1983; WHO, 1985).
Because the human body can adapt to low and high intakes of nitrogen, there is a substantial difference between intakes barely sufficient to compensate for losses or to permit growth and intakes that may be associated with harmful effects. Since there are few criteria by which to evaluate the significance of the rate of protein turnover and pool size, value judgments must be made as to what is desirable in adults. For children, the protein required for growth is relatively small compared to that needed for maintenance. Nevertheless, satisfactory growth is a sensitive indicator of protein nutritional status.
The requirement for protein is reasonably well established for the very young child and the young male adult. For other age groups, much less information is available, and protein needs are estimated in part by interpolation or extrapolation based on reasonable biological principles.
THE REQUIREMENT FOR AMINO ACIDS
In determining the requirement for protein, the subcommittee first considered requirements for the essential amino acids. The required amounts of the nine essential amino acids must be provided in the diet, but because cystine can replace approximately 30% of the requirement for methionine, and tyrosine about 50% of the requirement for phenylalanine, these amino acids must also be considered. The essential amino acid requirements of infants, children, men, and women were studied extensively from 1950 to 1970. Except for infants, where the criterion was growth and nitrogen accretion, the requirement was accepted to be the amount of intake needed to achieve nitrogen equilibrium in short-term studies of adults or positive balance in children (see review by FAO/WHO, 1973; NRC, 1974; WHO, 1985). Estimates of amino acid requirements for various age groups are listed in Table 6-1.
TABLE 6-1
Estimates of Amino Acid Requirements.
In a novel approach to examining these requirements, the need for four amino acids was examined in children whose diets were strictly controlled because of inborn errors of metabolism and who were developing normally (Kindt and Halvorsen, 1980). Requirements determined in this way during the first 3 years of life are in good agreement with the values for isoleucine, leucine, phenylalanine plus tyrosine, and valine given in Table 6-1 for infants and 2-year-old children.
The requirement for histidine has not been quantified beyond infancy. Requirement values are difficult to establish because deficiency symptoms occur only after long periods of low intake. Kopple and Swendseid (1981) demonstrated that nitrogen balance diminished when histidine intake was less than 2 mg/kg per day, and increased when intake was increased to 4 mg/kg per day. WHO (1985) estimated the probable adult histidine requirement to be between 8 and 12 mg/kg per day by extrapolation from the infant requirement; this estimate is likely to be high, but safe.
The relatively low requirements estimated for adults have been confirmed by Inoue et al. (1988) using the nitrogen balance method. Studies of whole body lysine, leucine, valine, and threonine oxidation rates suggest that adult requirements for these essential amino acids have been underestimated. Approximations of average requirements according to the 13C tracer studies are leucine, 40 mg/kg (Meguid et al., 1986a); lysine, 35 mg/kg (Meredith et al., 1986); threonine, 15 mg/kg (Zhao et al., 1986); and valine, 16 mg/kg (Meguid et al., 1986b). These new estimates have been challenged on methodologic and theoretical grounds (Millward and Rivers, 1986) and require further confirmation.
Studies on requirements for individual essential amino acids in the elderly are inconsistent. Some suggest that requirements are increased in the elderly; others indicate that they are decreased (Munro, 1983). In the one study in which the same methodology and design were applied to the elderly as in a study of young men, no differences in requirements between age groups were found (Watts et al., 1964). The pattern of requirement for essential amino acids in the elderly is accepted to be the same as for younger adults.
There is no information on amino acid requirements of pregnant and lactating women.
The data demonstrate the unsatisfactory state of knowledge concerning amino acid requirements. The values in Table 6-1 are the best available and serve as the basis for calculation of amino acid requirement patterns at various ages and for procedures for the amino acid scoring of diets (see below).
RECOMMENDED ALLOWANCES FOR PROTEIN
In establishing an RDA for protein, three steps were followed: (1) The subcommittee first estimated the average requirement for reference proteins (i.e., highly digestible, high-quality protein such as egg, meat, milk, or fish) according to sex, age, and reproductive status of women. (2) The standard deviation of requirement was determined and average requirement values were increased accordingly to compute the recommended allowance of reference protein. (3) Amino acid scoring patterns were tabulated. These were based on requirements of various age groups for essential amino acids and for total protein. These patterns of requirement were reviewed in relation to U.S. food consumption patterns to determine if adjustment of the allowance for reference protein would be warranted in establishing the RDA for protein due to amino acid composition or protein digestibility of food proteins consumed.
The Requirement for Reference Protein and Its Variability
Adults To determine the protein requirements of young male adults, WHO (1985) reviewed evidence from both short- and longterm nitrogen balance studies. On the basis of recalculated data from the short-term studies, the international group proposed a mean requirement of 0.61 g/kg per day for reference protein. Several relatively long-term studies (58 to 89 days) involving single levels of protein intake yielded similar estimates of requirement for subjects consuming egg-protein diets. By averaging the two sets of balance data (i.e., from the long- and short-term studies), a protein requirement of 0.6 g/kg per day (rounded) was obtained. This is accepted to be the average daily requirement for reference proteins. No data were available on the coefficient of variation for long-term studies, but for short-term studies, the mean coefficient of variation was estimated to be 12.5%. A value of 25% (2 SDs) above the average requirement would be expected to meet the needs of 97.5% of a normally distributed population. Thus, 0.75 g/kg per day (0.6 × 1.25) is the recommended allowance of reference protein for young male adults.
The international group examined the data from several short-term studies in which men were fed habitual mixed diets of ordinary food. The requirements were predicted to be 0.54 to 0.99 g/kg per day, the larger estimates deriving from diets of lower digestibility and quality. The adult requirement for absorbed protein appears not to differ between reference and practical diets.
There are fewer data for young adult women, but there is evidence (Calloway and Kurzer, 1982) that requirement values, when adjusted for body weight, are not substantially different from those for young adult men. Accordingly, the recommended allowance for reference protein is 0.75 g/kg per day for both sexes.
The Elderly The protein content of the adult body diminishes with age. More specifically, nonmuscle mass is little affected by age, whereas muscle diminishes extensively and is compensated for by an increase in body fat. These changes in muscle mass are related to whole-body protein turnover and changes in the rate of protein synthesis (Uauy et al., 1978a). Muscle protein turnover accounts for 30% of the total protein turnover in the young adult, but only 20% of that in the elderly (Munro, 1983). Serum albumin levels and daily albumin synthesis also decrease in elderly people who consume diets with adequate protein (Gersovitz et al., 1980).
Questions thus arise concerning the extent to which changes in protein metabolism affect protein requirements. Dietary protein needs might be expected to change during the aging process, i.e., to decrease due to lower rates of turnover, or to increase due to decreased efficiency of absorptive and metabolic processes and to decreased total food intake associated with reduced physical activity, or to become more variable due to disease and disability. There is surprisingly little information on which to base recommendations for protein intake in the elderly.
The early literature on this subject was reviewed by Irwin and Hegsted (1971). Only a few studies have been conducted in the more recent past, and their results are inconsistent. Zanni et al. (1979) concluded that 0.57 g of egg protein per kilogram of observed body weight (weights were above ideal for height) was adequate to sustain nitrogen equilibrium at a food energy intake of 30 kcal/kg, whereas Uauy et al. (1978b) found that 0.57 g/kg was insufficient for nearly all subjects and that needs were only barely met at 0.8 g/kg. According to Gersovitz et al. (1982), 0.8 g of egg protein per kilogram of body weight per day was insufficient to maintain nitrogen balance in the majority of elderly men and women studied over a 30-day period, but Cheng et al. (1978) found this level of protein to be adequate. In both of these studies, body weight was maintained; however, energy intake was lower in the study by Gersovitz and colleagues (1982), suggesting that activity patterns may have been different in the two groups or that nitrogen balance was improved by the higher food energy intake in the study by Cheng et al. (1978). Variations in activity level, disease prevalence, and use of therapeutic drugs are all potentially confounding variables.
The recommended allowance for reference protein (0.75 g/kg) is accepted to be the same for the elderly as for young adults. Because of the difference in body composition, this allowance is higher per unit of lean body mass and should allow for some decrement in utilization efficiency.
Pregnancy Additional protein is required during pregnancy for the mother and the fetus (Hytten and Leitch, 1971). Maternal protein synthesis increases in order to support expansion of the blood volume, uterus, and breasts, and fetal and placental proteins are synthesized from amino acids supplied by the mother. The magnitude of the required increase in dietary intake remains uncertain, since different methods of estimation yield different figures.
The factorial method of estimating requirements for pregnancy is based on the amount of protein present in the fetus, placenta, and maternal tissues, including blood (Hytten and Leitch, 1971). It is calculated that 925 g of protein is deposited during a pregnancy involving 12.5 kg of maternal weight gain and an infant weighing 3.3 kg at term. The rate of nitrogen retention is not constant; for the first through third trimester, nitrogen deposition is estimated to be 0.11, 0.52, and 0.92 g/day, respectively (Table 6-2). Evidence from animal studies suggests, however, that relatively more protein may be stored during early gestation and mobilized at later stages of pregnancy (Naismith, 1977; Naismith and Morgan, 1976). Protein turnover is increased by the twelfth week of pregnancy (de Benoist et al., 1985); it remains high in the second trimester and the same (Jackson, 1987) or lower (Fitch and King, 1987) in the third trimester. Consequently, increased protein needs during pregnancy may be more uniform across time than the figures of Hytten and Leitch indicate.
TABLE 6-2
Derivation of Reference Protein Allowance During Pregnancy.
Nitrogen balance data can also be used to estimate protein requirements during pregnancy. Almost all older balance studies have indicated that more nitrogen is retained during pregnancy than is predicted on the basis of fetal and placental growth and maternal tissue hypertrophy (Calloway, 1974). In more recent studies (Appel and King, 1979; Johnstone et al., 1981), nitrogen retentions have been found to be closer to the theoretical value if allowance is made for unmeasured losses. Nevertheless, data on changes in body weight and body potassium indicate that nitrogen retention under laboratory conditions is somewhat greater than can be accounted for only by the fetus and maternal supportive tissue (Appel and King, 1979; King et al., 1973). This suggests that protein might be retained at sites other than those now recognized, for example, in skeletal muscle. This possibility is supported in part by analyses of animal carcasses (King, 1975).
Dietary surveys in developed countries indicate that pregnant women eating self-selected diets generally consume somewhat larger amounts of protein than theoretical requirements. Moreover, satisfactory levels of protein intake tend to be associated with improved reproductive outcome (Higgins et al., 1973; Lechtig et al., 1975; Metcoff et al., 1981; Osofsky, 1975). However, such epidemiological data are confounded by the strong dietary correlation between protein and energy, as well as their metabolic interrelationships.
Despite these discrepancies between factorial and nitrogen balance estimates of nitrogen gain during pregnancy, the subcommittee concurs with WHO (1985) that the estimate of requirement should be based on the factorial method. The coefficient of variation in birth weight is 15%—a variance assigned to all components of pregnancy protein gain. The average storage value is thus increased by 30% to include the protein gains of virtually all healthy women during pregnancy. These values must be adjusted for the efficiency with which dietary protein is converted to fetal, placental, and maternal tissues; this is generally assumed to be 70%—the same as efficiency observed in infants. The additional allowance of reference protein needed to support the deposition of new tissue is calculated to be 1.3, 6.1, and 10.7 g/day during the first, second, and third trimesters of pregnancy, respectively (Table 6-2). There is also a maintenance requirement associated with the added lean tissue. To allow for this and because of the uncertainty about the rate of tissue deposition, the subcommittee recommends an additional allowance of 10 g of reference protein per day throughout pregnancy.
Lactation The average protein (nitrogen × 6.25) content of mature human milk is approximately 1.1 g/100 ml, except during the first month when it is about 1.3 g/100 ml (WHO, 1985). Analyses of human milk composition in the United States, based on a study of 40 mothers in the first 4 months of lactation, show a fall in protein content from 1.36 g/100 ml to 1.12 g/100 ml during this period (Butte et al., 1984).
The average protein requirement for lactation is estimated from milk composition and the mean volume of milk produced, which is 750 ml/day, adjusted for 70% efficiency in the conversion of dietary protein to milk protein. The coefficient of variation is taken to be 12.5% (WHO, 1985); thus, the average requirement is increased by 25% to determine the recommended allowance of a reference protein. The allowance is calculated as follows:

The volume of milk produced in the second 6 months of lactation is about 20% less than in the first 6 months, according to international data (WHO, 1985). No recent data from the United States have been published. If the international figure is accepted, then the additional allowance in this period is 11.8 g of reference protein per day. The recommended allowance of additional reference protein is 15 g/day during the first 6 months of lactation and 12 g/day thereafter.
Infants, Children, and Adolescents During the first year of life, the protein content of the body increases from 11 to 15%, and body weight increases by approximately 7 kg. The average increase in body protein is about 3.5 g/day during the first 4 months of life and 3.1 g/day during the subsequent 8 months (Fomon, 1974). By 4 years of age, body protein content reaches the adult value of 18 to 19% of body weight (Widdowson and Dickerson, 1963). As the growth rate drops rapidly after the first year of life, the maintenance requirement represents a gradually increasing proportion of the total protein requirement.
For the first months of life, requirements are based on intake data because of the difficulty in accurately estimating allowances for growth and maturation of body composition. Infants breastfed by healthy, well-nourished mothers or fed human milk by bottle can grow at a satisfactory rate for about 4 months (Butte et al., 1984; Ferris et al., 1978; Fomon, 1986). Protein d intakes by breastfed infants range from 2.43 g/kg per day in the first month to 1.51 g/kg per day in the fourth month, averaging 2.04 g/kg per day in the first 3 months and 1.73 g/kg per day in the next 3 months (WHO, 1985). Breastfed infants in the United States grow satisfactorily at a mean protein intake of 1.68 g/kg per day (total nitrogen × 6.25) during the first 3 months (Butte et al., 1984). Probability assessment suggests that the true requirement appears to be less than mean intake; the figure proposed is 1.1 g/kg with a deviation of 0.1 to 0.2 g/kg (Beaton and Chery, 1988). The subcommittee has, however, accepted an average intake of 1.68 g/kg per day as the requirement between birth and 3 months of age, the reference being human milk protein. The protein needs of an infant up to 4 months of age will be met if the energy needs are met, provided the food is human milk or a formula that contains protein of a quality and quantity equivalent to that of human milk.
A modified factorial procedure for calculating the protein needs of infants and children was examined by the WHO group. Values were assigned for a maintenance requirement, and an increment representing growth, the theoretical value for which was increased by 50% to allow for marked unevenness of daily growth rate and the inability of infants and children to store amino acids against intermittent needs. Efficiency of utilization was accepted to be 70%. These estimates were compared with data from the few studies of nitrogen balance in the age group 6 months to 9 years. Neither the factorial nor balance estimates were consistently higher or lower across age groups. Because there are large gaps in the experimental data, the modified factorial approach was used by the WHO group to estimate needs for all children. The international values are accepted by this subcommittee in establishing average requirements for reference protein beyond age 3 months. Table 6-3 provides examples of the steps involved in the calculations of daily allowances by the modified factorial approach. The coefficient of variation in requirements calculated in this manner is assumed to be 12.5%—the same as that established for maintenance requirements in adults. The factorially determined average requirement is accordingly increased by 25% to derive the recommended allowance for reference protein.
TABLE 6-3
Examples of the Derivation of Protein Allowances for Children and Adolescents by a Factorial Procedure.
The tabulated values for reference protein allowances for the various age and sex groups are listed in Table 6-4. To convert these values to daily allowances of average U.S. dietary protein, one must examine these values in relation to the amino acid composition and digestibility of proteins consumed. Since the requirements of both amino acids and protein differ among age and sex groups, the quality of protein required to meet these needs will also vary.
TABLE 6-4
Recommended Allowances of Reference Protein and U.S. Dietary Protein.
Amino Acid Requirement Pattern According to Age
A pattern of requirements for amino acids in the total dietary protein is calculated by dividing each essential amino acid requirement by the recommended allowance of reference protein for the given age group (see Table 6-5).
TABLE 6-5
Amino Acid Requirement PatternsCompared with the Composition of High-Quality Proteinsand the U.S. Diet.
The requirement pattern for infants is based on quantitative amino acid requirements (Table 6-1) divided by the reference protein allowance of infants 3 to 4 months of age (1.73 g/kg; Table 6-3)—the age at which the studies were made. WHO (1985) accepted for this age group a pattern based on the average composition of human milk protein. Both patterns are shown in Table 6-5. The variation in reported composition of human milk proteins is large, and a substantial portion of breast milk nitrogen is nonprotein. Given the difficulties in estimating the factors that affect the patterns, the values are in surprisingly good agreement. Only for tryptophan is the difference substantial. This subcommittee concludes that the composition of human milk should be used as a reference pattern for formulation of human milk substitutes for infants and as a guide to supplementary feeding of infants through 6 months of age.
The amino acid pattern shown for 2-year-old children should be applied to children ages 2 to 6, and that shown for children 10 to 12 years should be used for ages 6 to 13 years. The adult pattern is applicable to children above age 13 and adults. For children above 6 months and less than 2 years of age, a combination of the infant and preschool child figures should be used to evaluate a total diet. These values and recommendations are in general agreement with those of WHO (1985).
There are no established amino acid requirements and, hence, no amino acid patterns for pregnancy and lactation. The international group suggested that a pattern for the lactating woman's total diet could be developed by summing the adult protein and additional lactation allowances, calculating the proportion of each in the total, and applying the adult pattern to its fractional part (0.72) and the human milk pattern to the other (0.28). Values derived in this way are, in mg/g of protein: lysine, 31; methionine and cystine, 21; threonine, 19; and tryptophan, 9. A similar calculation is theoretically possible for pregnancy, but more difficult because of the variety of tissues and proteins deposited.
Amino Acid Scoring of Dietary Protein Quality To adjust for amino acid composition, a score is calculated according to the most limiting amino acid, i.e., the one in greatest deficit for the age group involved.

The amino acid score should be based on the appropriate pattern for age. Only four essential amino acids are likely to affect the protein quality of mixed human diets: lysine, the sulfur-containing amino acids (methionine plus cystine), threonine, and tryptophan. Scoring patterns for all the essential amino acids (presented in Table 6-5) are important for the formulation of special purpose diets in clinical practice.
CONSUMPTION AND AMINO ACID PATTERN OF PROTEINS IN THE U.S. DIET
Food consumption data from the U.S. Department of Agriculture's (USDA) 1977–1978 and 1985 surveys indicate that 14 to 18% of the total food energy intake is derived from protein (USDA, 1983, 1986, 1987). Despite wide variations in food energy intake, this proportion remains similar for both sexes and all age groups except infants. There is also little change as a function of household income, urbanization, or race. Food items likely to be underreported in surveys (e.g., alcoholic beverages, confections) would provide energy but little protein; hence, the percentage of energy from protein may be overestimated. Average consumption levels are, however, quite generous: about 50 g/day in preschool children; 70 to 85 g in older children; 90 to 110 g in male and 65 to 70 g in female adolescents and adults; and 75 to 80 g in men and 55 to 65 g in women over age 65.
Foods of animal origin contribute approximately 65% of the protein in the USDA survey, with the proportion from the meat and dairy groups varying somewhat with age (USDA, 1983). Similarly, the data from the second National Health and Nutrition Examination Survey (NHANES II) indicate that about 48% of the protein is derived from meat, fish, and poultry; 17% from dairy products; and 4% from eggs (Block et al., 1985). The importance of grain products as suppliers of protein is not always realized, particularly in populations ingesting diets rich in animal products. Cereal grains supply an average of 16 to 20% of the total protein intake in the United States.
The amino acid pattern in the diet consumed by children ages 1 to 3 years and all persons surveyed is given in Table 6-5. The pattern is uniform between the age groups and meets the requirement pattern levels for all age groups except infants. The U.S. consumption pattern also meets the provisional pattern for lactating women. Therefore, no adjustment to the recommended allowance for reference protein is required for people consuming a typical U.S. diet.
Digestibility The amino acid score alone may lead to an overestimation of the capacity of some proteins to meet physiological requirements unless digestibility is taken into account. When the amino acid score is multiplied by digestibility, it becomes analogous to the biologically determined net protein utilization (NPU). The NPU is the product of biological value (comparable to amino acid score) and true protein digestibility.
Differences in digestibility result from intrinsic differences in the nature of food protein and the nature of the cell wall, from the presence of other dietary factors that modify digestion (e.g., dietary fiber, polyphenols such as tannins, and enzyme inhibitors), and from chemical reactions (e.g., binding of the amino groups of lysine and cross-linkages), which may affect the release of amino acids by enzymatic processes. There are few data on the digestibility of specific amino acids in food proteins, and any differences are not captured in measurements of overall protein digestibility. Although it is known that there are differences between the pattern of amino acids in food protein, fecal matter, and portal blood, it is not now possible to provide finer adjustment than overall digestibility.
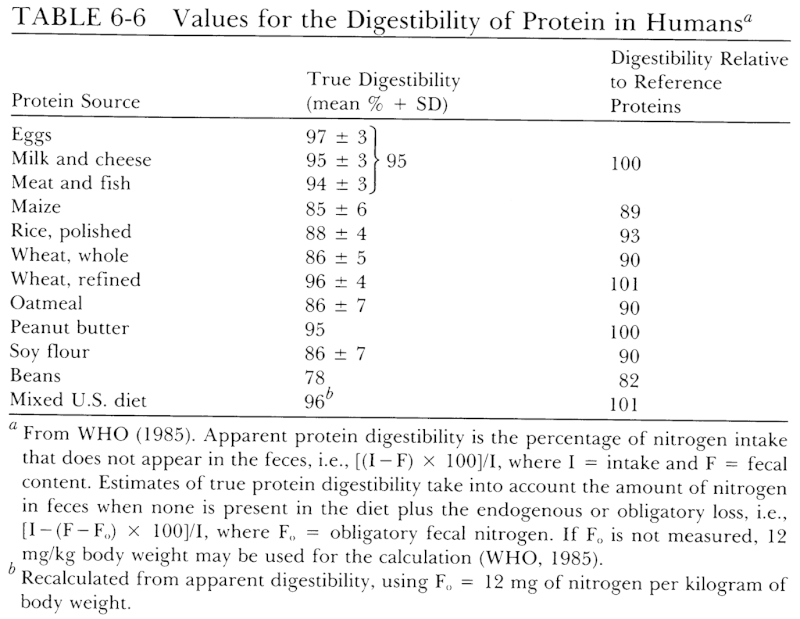
Representative data on the digestibility of some selected proteins are shown in Table 6-6. A more comprehensive listing of protein digestibility can be found in reports by Hopkins (1981) and FAO (1970). The true digestibility of reference proteins is assigned a value of 100 for translating requirements for reference proteins to recommended levels of intake for ordinary mixtures of dietary proteins. Since the mixed protein of a typical U.S. diet is shown to be as well digested as reference proteins, no adjustment for this factor is normally required.
Adjustment of Allowances for Dietary Quality Adjustment for exceptional dietary patterns can be made by deriving a weighted digestibility factor based on the digestibilities of the principal protein sources consumed and an amino acid score based on their contribution of essential amino acids. Such adjustment would rarely be warranted for the U.S. population. Shown in Table 6-7 is an example of calculations required to make an adjustment for an unusual diet—one in which the usual consumption pattern is reversed, i.e., only one-third of the protein from animal sources. A comparison of the amino acid pattern with the requirement patterns in Table 6-5 shows that lysine is low for the preschool age group and tryptophan is borderline. The limiting amino acid is lysine, which has a score of 51/58, or 88%. The amino acid pattern meets the requirement patterns of older children and adults, i.e., the score is 100. The weighted digestibility factor is 92%. Thus, the protein allowance for a 3-year-old child is 1.1 × 100/88 × 100/92, or 1.4 g/kg. For older children and adults, an adjustment of the allowance would be made only for digestibility.
TABLE 6-7
Example of Calculations Needed for Adjustment of Protein Allowances for a Diet with 33% Animal- and 67% Vegetable-Source Protein.
OTHER EFFECTS ON PROTEIN REQUIREMENTS
There is little evidence that muscular activity increases the need for protein, except for the small amount required for the development of muscles during physical conditioning (Torun et al., 1977). Vigorous activity that leads to profuse sweating, such as in heavy work and sports, and exposure to heat increases nitrogen loss from the skin, but with acclimatization to a warm environment, the excessive skin loss is reduced and may be partially compensated by decreased renal excretion (WHO, 1985). In view of the margin of safety in the RDA, no increment is added for work or training.
No added allowance is made here for the usual stresses encountered in daily living, which can give rise to transient increases in urinary nitrogen output (Scrimshaw et al., 1966). It is assumed that the subjects of experiments forming the basis for the requirement estimates are usually exposed to the same stresses as the population generally.
Extreme environmental or physiological stress increases nitrogen loss (Cuthbertson, 1964). Infections, fevers, and surgical trauma can result in substantial nitrogen loss through the urine and greatly increased energy expenditure. Therefore, severe infections and surgery should be treated as clinical conditions that require special dietary treatment. During convalescence from an illness that has led to protein depletion, requirements for both protein and energy are elevated because of the need to replace wasted tissues, just as they are during periods of rapid growth. Premature infants also require special consideration with regard to amino acid composition of the formulas and level of protein intake.
COMPARISON OF CURRENT PROTEIN RECOMMENDATIONS TO THE 1980 RDAS
The RDAs for protein are summarized in Table 6-4. After rounding, allowances are (in g/kg per day): children 1 to 3 years old, 1.2; 4- to 6-year-old children, 1.1; 7- to 14-year-old children, 1.0; 15- to 18-year-old boys, 0.9; and all others (except infants), 0.8. The 1980 RDA for infants was 2.2 g/kg during the first 6 months and 2.0 g/kg for those between 6 and 12 months of age. The present RDA for the 6- to 12-month age group is somewhat lower, having been based on newer data on observed intakes of healthy children and theoretical growth requirements. Human milk or an equivalent substitute (i.e., formula containing as much protein and a similar amino acid pattern, corrected for digestibility if warranted) is recommended for infants from birth to 3 months of age. RDAs for other age and sex groups are essentially the same as the 1980 figures, but the derivation and justification are different, reflecting both new data and reexamination of older data. Present allowances for pregnancy and lactation are lower than in 1980: 10 g/day in contrast to 30 g/day in pregnancy, and 15 g/day rather than 20 g/day in lactation. The revised allowance for pregnancy is more heavily influenced by theory than by new evidence; the allowance for lactation is in accord with new information on breast milk production by women in the United States.
EXCESSIVE INTAKES AND TOXICITY
Because the system for disposal of excess nitrogen is efficient, protein intakes moderately above requirement are believed to be safe. Brenner et al. (1982) postulated that excess protein intake accelerates the processes that lead to renal glomerular sclerosis, a common phenomenon of aging. There is supportive evidence from studies in animals, but not in humans on this point. Urinary calcium excretion increases with increased protein intake if phosphorus intake is constant. If phosphorus intake increases with protein intake, as it does in U.S. diets, the effect of protein is minimized (Hegsted et al., 1981; Schuette and Linkswiler, 1982). It has been suggested, but not demonstrated, that a habitual high intake of protein might contribute to osteoporosis. This seems unlikely based on present evidence, at least for the range of intake by most people in the United States. Habitual intakes of protein in the United States are substantially above the requirement, and although there is no firm evidence that these intake levels are harmful, it has been deemed prudent to maintain an upper bound of no more than twice the RDA for protein (NRC, 1989).
REFERENCES
- Appel, J., and J.C. King. 1979. Protein utilization in pregnant and nonpregnant women. Fed. Proc. 38: 388.
- Beaton, G.H., and A. Chery. 1988. Protein requirements of infants: a reexamination of concepts and approaches. Am. J. Clin. Nutr. 48: 1403–1412. [PubMed: 3202089]
- Block, G., C.M. Dresser, A.M. Hartman, and M.D. Carroll. 1985. Nutrient sources in the American diet: quantitative data from the NHANES II survey. II. Macronutrients and fats. Am. J. Epidemiol. 122: 27–40. [PubMed: 4014199]
- Brenner, B.M., T.W. Meyer, and T.H. Hostetter. 1982. Dietary protein intake and the progressive nature of kidney disease: the role of hemodynamically mediated glomerular injury in the pathogenesis of progressive glomerular sclerosis in aging, renal ablation, and intrinsic renal disease. N. Engl. J. Med. 307: 652–659. [PubMed: 7050706]
- Butte, N.F., C. Garza, E.O. Smith, and B.L. Nichols. 1984. Human milk intake and growth in exclusively breast-fed infants. J. Pediatr. 104: 187–195. [PubMed: 6694010]
- Calloway, D.H. 1974. Nitrogen balance during pregnancy. Pp. 79–94 in M. Winick, editor. , ed. Nutrition and Fetal Development. John Wiley & Sons, New York.
- Calloway, D.H., and M.S. Kurzer. 1982. Menstrual cycle and protein requirements of women. J. Nutr. 112: 356–366. [PubMed: 7057271]
- Cheng, A.H.R., A. Gomez, J.G. Bergan, T.C. Lee, F. Monckeberg, and C.O. Chichester. 1978. Comparative nitrogen balance study between young and aged adults using three levels of protein intake from a combination wheat-soy-milk mixture. Am. J. Clin. Nutr. 31: 12–22. [PubMed: 579565]
- Cho, E.S., H.L. Anderson, R.L. Wixom, K.C. Hanson, and G.F. Krause. 1984. Long-term effects of low histidine intake on men. J. Nutr. 114: 369–384. [PubMed: 6693997]
- Cuthbertson, D.P. 1964. Physical injury and its effects on protein metabolism. Pp. 374–414 in H.N. Munro, editor; and J.B. Allison, editor. , eds. Mammalian protein Metabolism, Vol II. Academic Press, New York.
- de Benoist, B., A.A. Jackson, J.S. Hall, and C. Persaud. 1985. Whole-body protein turnover in Jamaican women during normal pregnancy. Human Nutr. Clin. Nutr. 39: 167–179. [PubMed: 3874856]
- FAO (Food and Agriculture Organization). 1970. Amino-Acid Content of Foods and Biological Data on Proteins. Food Policy and Food Science Service, Nutrition Division. FAO Nutritional Studies No. 24. Food and Agriculture Organization, Rome. 285 pp.
- FAO/WHO (Food and Agriculture Organization/World Health Organization). 1973. Energy and Protein Requirements. Report of a Joint FAO/WHO Ad Hoc Expert Committee. Technical Report Series No. 552; FAO Nutrition Meetings Report Series 52. World Health Organization, Rome. 118 pp.
- Ferris, A.G., L.B. Vilhjalmsdottir, V.A. Beal, and P.L. Pellett. 1978. Diets in the first six months of infants in western Massachusetts. I. Energy-yielding nutrients. J. Am. Diet. Assoc. 72: 155–160. [PubMed: 624810]
- Fitch, W.L., and J.C. King. 1987. Protein turnover and 3-methylhistidine excretion in nonpregnant, pregnant and gestational diabetic women. Human Nutr. Clin. Nutr. 41C: 327–339. [PubMed: 3692907]
- Fomon, S.J. 1974. Infant Nutrition, 2nd ed. W.B. Saunders, Philadelphia. 575 pp.
- Fomon, S.J. 1986. Protein requirements of term infants. Pp. 55–68 in S.J. Fomon, editor; and W.C. Heiod, editor. , eds. Energy and Protein Needs During Infancy. Academic Press, New York.
- Fomon, S.J., and L.J. Filer, Jr. 1967. Amino acid requirements for normal growth. Pp. 391–401 in W.L. Nyhan, editor. , ed. Amino Acid Metabolism and Genetic Variation. McGraw-Hill, New York.
- Garza, C., N.S. Scrimshaw, and V.R. Young. 1977. Human protein requirements: a long-term metabolic nitrogen balance study in young men to evaluate the 1973 FAO/WHO safe level of egg protein intake. J. Nutr. 107: 335–352. [PubMed: 833693]
- Gersovitz, M., H.N. Munro, J. Udall, and V.R. Young. 1980. Albumin synthesis in young and elderly subjects using a new stable isotope methodology: response to level of protein intake. Metabolism 29: 1075–1086. [PubMed: 7432171]
- Gersovitz, M., K. Motil, H.N. Munro, N.S. Scrimshaw, and V.R. Young. 1982. Human protein requirements: assessment of the adequacy of the current Recommended Dietary Allowance for dietary protein in elderly men and women. Am. J. Clin. Nutr. 35: 6–14. [PubMed: 7064878]
- Hegsted, D.M. 1976. Balance studies. J. Nutr. 106: 307–311.
- Hegsted, D.M., S.A. Schuette, M.B. Zemel, and H.M. Linkswiler. 1981. Urinary calcium and calcium balance in young men as affected by level of protein and phosphorus intake. J. Nutr. 111: 553–562. [PubMed: 7205408]
- Heird, W.C., J.F. Nicholson, J.M. Driscoll, Jr., N.J. Schullinger, and R.W. Winters. 1972. Hyperammonemia resulting from intravenous alimentation using a mixture of synthetic L-amino acids: a preliminary report. J. Pediatr. 81: 162–165. [PubMed: 4624624]
- Higgins, A.C., E.W. Crampton, and J.E. Moxley. 1973. Nutrition and the outcome of pregnancy. Pp. 1071–1077 in R.O. Scow, editor. , ed. Endocrinology. Proceedings of the Fourth International Congress of Endocrinology, Washington, D.C., June 18–24, 1972. Excerpta Medica, Amsterdam.
- Hopkins, D.T. 1981. Effects of variation in protein digestibility. Pp. 169–193 in C.E. Bodwell, editor; , J.S. Adkins, editor; , and D.T. Hopkins, editor. , eds. Protein Quality in Humans: Assessment and In Vitro Estimation. AVI Publishing, Westport, Conn.
- Horowitz, J.H., E.B. Rypins, J.M. Henderson, S.B. Heymsfield, S.D. Moffitt, R.P. Bain, R.K. Chawla, J.C. Bleier, and D. Rudman. 1981. Evidence for impairment of transsulfuration pathway in cirrhosis. Gastroenterology 81: 668–675. [PubMed: 7262512]
- Hytten, F.E., and I. Leitch. 1971. The Physiology of Human Pregnancy, 2nd ed. Blackwell Scientific Publications, Oxford. 599 pp.
- Inoue, G., T. Komatsu, K. Kishi, and Y. Fujita. 1988. Amino acid requirements of Japanese young men. Pp. 55–62 in G.L. Blackburn, editor; , J.P. Grant, editor; , and V.R. Young, editor. , eds. Amino Acids: Metabolism and Medical Applications. John Wright, Boston.
- Irwin, M.I., and D.M. Hegsted. 1971. A conspectus of research on protein requirements of man. J. Nutr. 101: 387–429. [PubMed: 5101676]
- Jackson, A.A. 1987. Measurement of protein turnover during pregnancy. Human Nutr. Clin. Nutr. 41C: 497–498. [PubMed: 3429269]
- Johnstone, F.D., D.M. Campbell, and I. MacGillivray. 1981. Nitrogen balance studies in human pregnacy. J. Nutr. 111: 1884–1893. [PubMed: 7299489]
- Kindt, E., and S. Halvorsen. 1980. The need of essential amino acids in children. An evaluation based on the intake of phenylalanine, tyrosine, leucine, isoleucine, and valine in children with phenylketonuria, tyrosine amino transferase defect, and maple syrup urine disease. Am. J. Clin. Nutr. 33: 279–286. [PubMed: 6101930]
- King, J.C., D.H. Calloway, and S. Margen. 1973. Nitrogen retention, total body 40K and weight gain in teenage pregnant girls. J. Nutr. 103: 772–785. [PubMed: 4710089]
- Kopple, J.D., and M.E. Swendseid. 1981. Effect of histidine intake on plasma and urine histidine levels, nitrogen balance and N T -methylhistidine excretion in normal and chronically uremic men. J. Nutr. 111: 931–942. [PubMed: 7241233]
- Lechtig, A., J.P. Habicht, H. Delgado, R.E. Klein, C. Yarbrough, and R. Martorell. 1975. Effect of food supplementation during pregnancy on birthweight. Pediatrics 56: 508–520. [PubMed: 1165955]
- Meguid, M.M., D.E. Matthews, D.M. Bier, C.N. Meredith, J.S. Soeldner, and V.R. Young. 1986. a. Leucine kinetics at graded leucine intakes in young men. Am. J. Clin. Nutr. 43: 770–780. [PubMed: 3518394]
- Meguid, M.M., D.E. Matthews, D.M. Bier, C.N. Meredith, and V.R. Young. 1986. b. Valine kinetics at graded valine intakes in young men. Am. J. Clin. Nutr. 43: 781–786. [PubMed: 3706188]
- Meredith, C.N., Z.M. Wen, D.M. Bier, D.E. Matthews, and V.R. Young. 1986. Lysine kinetics at graded lysine intakes in young men. Am. J. Clin. Nutr. 43: 787–794. [PubMed: 3085468]
- Metcoff, J., editor; , E.R. Klein, editor; and B.L. Nichols, editor. , eds. 1981. Workshop on nutrition of the child: maternal nutritional status and fetal outcome. Am. J. Clin. Nutr. 34: 653–817. [PubMed: 7223680]
- Millward, D.J., and J.P.W. Rivers. 1986. Protein and amino acid requirements in the adult human. J. Nutr. 116: 2559–2561. [PubMed: 3806249]
- Munro, H.N. 1983. Protein nutriture and requirement in elderly people. Bibl. Nutr. Dieta 33: 61–74. [PubMed: 6360153]
- Naismith, D.J. 1977. Protein metabolism during pregnancy. Pp. 503–511 in E.E. Philipp, editor; , J. Barnes, editor; , and M. Newton, editor. , eds. Scientific Foundations of Obstetrics and Gynaecology, 2nd ed. Year Book Medical Publishers, Chicago.
- Naismith, D.J., and B.L. Morgan. 1976. The biphasic nature of protein metabolism during pregnancy in the rat. Br. J. Nutr. 36: 563–566. [PubMed: 1009078]
- Nakagawa, I., T. Takahashi, T. Suzuki, and K. Kobayashi. 1964. Amino acid requirements of children: nitrogen balance at the minimum level of essential amino acids. J. Nutr. 83: 115–118. [PubMed: 14167700]
- NRC (National Research Council). 1974. Improvement of Protein Nutriture. Report of the Committee on Amino Acids, Food and Nutrition Board. National Academy of Sciences, Washington, D.C. 201 pp.
- NRC (National Research Council). 1989. Diet and Health: Implications for Reducing Chronic Disease Risk. Report of the Committee on Diet and Health, Food and Nutrition Board. National Academy Press, Washington, D.C. 750 pp. [PubMed: 25032333]
- Osofsky, H.J. 1975. Relationships between nutrition during pregnancy and subsequent infant and child development. Obstet. Gynecol. Surv. 30: 227–241. [PubMed: 804688]
- Pineda, O., B. Torun, F.E. Viteri, and G. Arroyave. 1981. Protein quality in relation to estimates of essential amino acids requirements. Pp. 29–42 in C.E. Bodwell, editor; , J.S. Adkins, editor; , and D.T. Hopkins, editor. , eds. Protein Quality in Humans: Assessment and In Vitro Estimation. AVI Publishing, Westport, Conn.
- Rand, W.M., N.S. Scrimshaw, and V.R. Young. 1981. Conventional (“long-term”) nitrogen balance studies for protein quality evaluation in adults: rationale and limitations. Pp. 61–94 in C.E. Bodwell, editor; , J.S. Adkins, editor; , and D.T. Hopkins, editor. , eds. Protein Quality in Humans: Assessment and In Vitro Estimation. AVI Publishing, Westport, Conn.
- Schuette, S.A., and H.M. Linkswiler. 1982. Effects on Ca and P metabolism in humans by adding meat, meat plus milk, or purified proteins plus Ca and P to a low protein diet. J. Nutr. 112: 338–349. [PubMed: 6276519]
- Scrimshaw, N.S., J.-P. Habicht, M.L. Piche, B. Cholakos, and G. Arroyave. 1966. Protein metabolism of young men during university examinations. Am. J. Clin. Nutr. 18: 321–324. [PubMed: 4951515]
- Solomons, N.W., and L.H. Allen. 1983. The functional assessment of nutritional status: principles, practice and potential. Nutr. Rev. 41: 33–50. [PubMed: 6346143]
- Torun, B., N.S. Scrimshaw, and V.R. Young. 1977. Effect of isometric exercises on body potassium and dietary protein requirements of young men. Am. J. Clin. Nutr. 30: 1983–1993. [PubMed: 412414]
- Uauy, R., J.C. Winterer, C. Bilmazes, L.N. Haverberg, N.S. Scrimshaw, H.N. Munro, and V.R. Young. 1978. a. The changing pattern of whole body protein metabolism in aging humans. J. Gerontol. 33: 663–671. [PubMed: 299555]
- Uauy, R., N.S. Scrimshaw, and V.R Young. 1978. b. Human protein requirements: nitrogen balance response to graded levels of egg protein in elderly men and women. Am. J. Clin. Nutr. 31: 779–785. [PubMed: 645626]
- USDA (U.S. Department of Agriculture). 1976–1989. USDA Agricultural Handbook Series 8. Foods—Raw, Processed, Prepared. Comparison of Nutrient Data Research. Human Nutrition Information Service. U.S. Department of Agriculture, Washington, D.C. (various pagings)
- USDA (U.S. Department of Agriculture). 1983. Nationwide Food Consumption Survey 1977–1978. Food Intakes: Individuals in 48 States, Year 1977–78. Report No. I-1. Consumer Nutrition Division, Human Nutrition Information Service. U.S. Department of Agriculture, Hyattsville, Md. 617 pp.
- USDA (U.S. Department of Agriculture). 1984. Nationwide Food Consumption Survey. Nutrient Intakes: Individuals in 48 States, Year 1977–78. Report No. 1-2. Consumer Nutrition Division, Human Nutrition Information Service. U.S. Department of Agriculture, Hyattsville, Md. 439 pp.
- USDA (U.S. Department of Agriculture). 1986. Nationwide Food Consumption Survey. Continuing Survey of Food Intakes by Individuals. Men 19–50 Years, 1 Day, 1985. Report No. 85-3. Nutrition Monitoring Division, Human Nutrition Information Service. U.S. Department of Agriculture, Hyattsville, Md. 94 pp.
- USDA (U.S. Department of Agriculture). 1987. Nationwide Food Consumption Survey. Continuing Survey of Food Intakes by Individuals: Women 19–50 Years and Their Children 1–5 Years, 4 Days, 1985. Report No. 85-4. Nutrition Monitoring Division, Human Nutrition Information Service, Hyattsville, Md. 182 pp.
- Watts, J.H., A.N. Mann, L. Bradley, and D.J. Thompson. 1964. Nitrogen balances of men over 65 fed the FAO and milk patterns of essential amino acids. J. Gerontol. 19: 370–374. [PubMed: 14179698]
- WHO (World Health Organization). 1985. Energy and Protein Requirements. Report of a Joint FAO/WHO/UNU Expert Consultation. Technical Report Series 724. World Health Organization, Geneva. 206 pp. [PubMed: 3937340]
- Widdowson, E.M., and J.W.T. Dickerson. 1963. Chemical composition of the body. Pp. 1–247 in C.L. Comar, editor; and F. Bronner, editor. , eds. Mineral Metabolism: An Advanced Treatise, Vol II. The Elements, Part A. Academic Press, New York.
- Williams, H.H., A.E. Harper, D.M. Hegsted, G. Arroyave, and L.E. Holt, Jr. 1974. Nitrogen and amino acid requirements. Pp. 23–63 in Improvement of Protein Nutriture. Report of the Committee on Amino Acids, Food and Nutrition Board. National Academy of Sciences, Washington, D.C.
- Young, V.R., and N.S. Scrimshaw. 1977. Human protein and amino acid metabolism and requirements in relation to protein quality. Pp. 11–54 in C.E. Bodwell, editor. , ed. Evaluation of Proteins for Humans. AVI Publishing, Westport, Conn.
- Zanni, E., D.H. Calloway, and A.Y. Zezulka. 1979. Protein requirements of elderly men. J. Nutr. 109: 513–524. [PubMed: 430255]
- Zhao, X.H., Z.M. Wen, C.N. Meredith, D.E. Matthews, D.M. Bier, and V.R. Young. 1986. Threonine kinetics at graded threonine intakes in young men. Am. J. Clin. Nutr. 43: 795–802. [PubMed: 3085469]
Footnotes
- a
In this chapter, protein is equated with nitrogen × 6.25, i.e., crude protein containing 16% nitrogen. Specific food proteins have greater (cereals) or lesser (milk) percentages of nitrogen. See USDA Agricultural Handbook Series 8 (1976–1989) for factors used in food composition tables.
- b
The WHO (1985) report contains extensive references to the original investigations. The reader is referred to that report for full documentation.
- c
The term protein requirement conventionally encompasses specific and nonspecific amino acid and amino nitrogen requirements.
- d
The figures given for protein are nitrogen × 6.25. Approximately 20% of the nitrogen in human milk is nonprotein nitrogen (e.g., amino acids, urea, nucleotides), much of which is assumed to be utilized by the infant, and by convention is calculated as if it were crude protein.
- GENERAL SIGNS OF DEFICIENCY
- GENERAL PRINCIPLES FOR ESTIMATING PROTEIN REQUIREMENTS
- OTHER CRITERIA OF ADEQUACY
- THE REQUIREMENT FOR AMINO ACIDS
- RECOMMENDED ALLOWANCES FOR PROTEIN
- CONSUMPTION AND AMINO ACID PATTERN OF PROTEINS IN THE U.S. DIET
- OTHER EFFECTS ON PROTEIN REQUIREMENTS
- COMPARISON OF CURRENT PROTEIN RECOMMENDATIONS TO THE 1980 RDAS
- EXCESSIVE INTAKES AND TOXICITY
- REFERENCES
- PubMedLinks to PubMed
- Protein and Amino Acids - Recommended Dietary AllowancesProtein and Amino Acids - Recommended Dietary Allowances
- CDKL1 cyclin dependent kinase like 1 [Homo sapiens]CDKL1 cyclin dependent kinase like 1 [Homo sapiens]Gene ID:8814Gene
Your browsing activity is empty.
Activity recording is turned off.
See more...