

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Feingold KR, Anawalt B, Blackman MR, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-.

ABSTRACT
Adrenal androgens (AA) are 19 carbon (C19) steroids that are secreted by the adrenal cortex through complicated biosynthetic pathways, which are regulated by complex mechanisms not completely understood as of yet. Adrenal steroidogenesis differs between the fetal and adult adrenal not only in regard to the site of production, but also in their significance for the human organism. The production of the AA is coordinated by a large number of adrenal and non-adrenal regulators. These steroids exert a number of effects in normal physiology and their excess may cause a number of different kinds of disorders.
INTRODUCTION
Adrenal androgens (AAs), normally secreted by the fetal adrenal zone and the zona reticularis of the adrenal cortex, are steroid hormones with weak androgenic activity. They include dehydroepiandrosterone (DHEA), dehydroepiandrosterone sulfate (DHEAS), androstenedione (A4), androstenediol (Α5) and 11β-hydroxyandrostenedione (11βOHA4) (1). DHEA and DHEAS are secreted in greater quantities than the other adrenal androgens. Although these steroids have little androgenic activity, they provide a pool of circulating precursors for peripheral conversion to more potent androgens (e.g. testosterone, T) and estrogens, (e.g. estradiol) (2-6). The production of T by the adrenal glands is minimal (7). Although adrenal androgens do not appear to play a major role in the fully androgenized adult man, they seem to play a role in the adult woman and in both sexes before puberty. Girls, women, and prepubertal boys may be negatively affected by AA hypersecretion in contrast to adult men. This chapter reviews AA biosynthesis, regulation, physiology and biological action. New data suggest that the principal androgen made by the human adrenal is 11-ketotestosterone (11-KT), a rarely studied steroid.
ADRENAL GLAND ANATOMY
Fetal Adrenal Gland

Figure 1:
Ontogenesis of steroidogenic enzymes in the human fetal adrenal gland. This schematic representation is divided into portions showing the fetal adrenal gland (right) at the first, second and third trimesters of pregnancy, and the adult adrenal gland (left). During the first trimester, the fetal gland is composed of a definitive zone (DZ, light grey) and a fetal zone (FZ, darker grey). Fetal zone (FZ) - expressing the P450C17 cytochrome, is responsible for massive secretion of DHEA and DHEA/S, used by the placenta as estrogen precursors. Second trimester - chromaffin cells (CC, darkest grey) originating from the neural crests migrate through the fetal cortex to progressively colonize the center of the gland to form the future medulla (Med). Third trimester - the newly constituted transitional zone (TZ, medium grey) acquires the enzyme 3ß-HSD while the expression of P450C17 remains, thus allowing the production of fetal cortisol. Near birth, cells of the definitive zone which express only 3ß-HSD, acquire the P450aldo and begin to secrete mineralocorticoids such as aldosterone. Neonatal - the fetal adrenal regresses strongly (mainly due to the regression of the fetal zone) and recovers progressively during the first years of extra-uterine life. Adult - adult adrenal gland is composed of the zona glomerulosa (ZGlo, light grey), zona fasciculata (ZFasc, medium grey) and zona reticularis (ZRet, darker grey) responsible for the production of mineralocorticoids (aldosterone), glucocorticoids (cortisol) and androgens (DHEA-DHEA/S), respectively. P450scc - cytochrome P450 side chain cleavage; Pregn. – pregnenolone; P450C17 - cytochrome P450 17a-hydroxylase, 17-20 lyase; 17OHP5 - 17-hydroxy-pregnenolone; DHEA/S - dehydroepiandrosterone-sulfate; S-Tfase - DHEA sulfotransferase; 3ß-HSD - 3ß-hydroxysteroid dehydrogenase; Prog. – progesterone; 17OHP4 - 17-hydroxyprogesterone; P450C21 - cytochrome P450 21-hydroxylase; P450C11 - cytochrome P450 11ß-hydroxylase; P450aldo - cytochrome P450 aldosterone synthase.
Fetal adrenal cortex arises from mesodermal cells migrating from the celomic epithelium very early in the embryonic period. Thus, adrenocortical tissue can be found in the ovaries, spermatic cord and testes. By the second month of gestation, the developing human fetal adrenal acquires two rudimentary, but distinct, zones: the inner fetal zone which consists of large eosinophilic cells, and the outer definitive zone, which is comprised of small, densely packed basophilic cells (8-9) (Figure 1). At about the ninth week of gestation, the developing human fetal adrenal is completely encapsulated. Ultrastructural studies also have revealed a third zone between the inner fetal zone and the definitive zone, the transitional zone (10). Cells in this zone show intermediate characteristics (11) and they demonstrate the capacity to synthesize cortisol, being histologically similar to cells of the zona fasciculata of the adult adrenal cortex. By the 30th week of gestation, the human fetal adrenal cortex manifests a rudimentary form of the adult adrenal cortex; the definitive zone and the transitional zone begin to resemble the zona glomerulosa and the zona fasciculata, respectively (12). Although the fetal zone is functionally similar to the adult zona reticularis (where DHEA-S is produced), it produces, unlike the adult zona reticularis, large amounts of other sulfated D5 steroids, including pregnenolone sulfate and 17a-hydroxypregnenolone sulfate.
Soon after birth, human fetal adrenal undergoes rapid involution due to the rapid regression of the inner fetal zone followed by a decrease in androgen secretion (12-17). Thus, the total weight of the glands decreases by approximately 50%. (11,18). Dramatic remodeling of the postnatal adrenal gland involves a complex combination of inner fetal zone regression and development of the zone glomerulosa and fasciculate (12,19). Because morphological studies have identified rudimentary zone glomerulosa and fasciculata during late gestation, the development of these zones may occur from their primordial structures, although there has been a general belief that the adult cortical zones develop from the persistent definitive zone (13).
Various genetic disorders of steroidogenesis, which constitute human “gene knockout experiments of nature”, indicate that fetal adrenal steroidogenesis, and the fetal adrenal zone itself, are not essential for fetal development, survival, or parturition (20). Aging results in tissue-rearrangements within the adrenal cortex while there is a relative increase of the outer cortical zones (21). As far it regards to the zona reticularis, after a continuous growth until young adulthood (20 to 25 years), it remains at a plateau for 5 to 10 years, and it regresses gradually after the reproductive period of life (22-23).
Adult Adrenal Gland
The adult adrenal glands, consisting of cortex and medulla, have a roughly pyramidal shape, lie above the upper poles of the kidneys in the retroperitoneum and weigh approximately 4g each. They are well supplied with arterial blood from branches of the phrenic arteries, the aorta, and the renal arteries, which give rise to the superior, middle, and inferior adrenal arteries, respectively. Arterial blood enters from the outer cortex, flows through fenestrated capillaries between the cords of cells, and drains into venules in the medulla. On the right side, the adrenal vein directly enters the inferior vena cava; on the left side, it usually drains into the left renal vein. The adrenal cortex is divided into three histologic and functional zones: the outer, aldosterone-secreting, called zona glomerulosa; the intermediate, predominantly cortisol and corticosterone secreting, called zona fasciculata; and the inner, predominantly androgens secreting called zona reticularis (Figure 2). Each one is characterized by the expression of specific steroidogenic enzymes, which result in the production of different steroid hormones. Zona glomerulosa constitutes about 15% of cortical volume. Zona fasciculata is the thickest part of the adrenal cortex, constructing about 75% of the cortex, produces cortisol as well as small amounts of androgens and estrogens. Zona reticularis surrounds the medulla and produces the adrenal androgens and small amounts of cortisol and estrogens (24). Zona glomerulosa is deficient in 17a – hydroxylase activity and thus cannot produce cortisol and androgens. Whereas zona glomerulosa is primarily regulated by angiotensin II and corticotropin (ACTH), both zona fasciculata and zona reticularis are regulated by ACTH (25). Both of these zones become hypofunctional and atrophic when ACTH is deficient while they become hypertrophic and hyperplastic when ACTH is secreted in excess.

Figure 2:
Schematic presentation of the adrenal zones and the main products of each zone. Downloaded from: georgiahealth.edu
The anatomical alterations of the adrenal cortex that occur during lifespan are followed by a marked decline in circulating adrenal C19 steroids and their resulting androgen metabolites. This decline takes place mainly between the age groups of 20-30 and 50-60 yr, with smaller changes observed after the age of 60 yr (26).
ADRENAL STEROIDS AND BIOSYNTHESIS OF ADRENAL ANDROGENS
Main Biosynthetic Pathway Of Adrenal Steroids
All human steroid hormones derive from cholesterol. Plasma lipoproteins are the major source of adrenal cholesterol. Low–density lipoprotein (LDL) accounts for about 80% of cholesterol delivered to the adrenal gland. There are specific cell surface LDL receptors on the adrenal tissue. Synthesis within the gland from acetyl-coenzyme A also occurs. A small pool of free cholesterol within the adrenal is available for acute response when stimulation occurs. Acute stimulation leads to hydrolysis of stored cholesteryl esters to free cholesterol, increased uptake from plasma lipoproteins and increased cholesterol synthesis within the gland (27). In addition, there is evidence that the adrenal can utilize high density lipoprotein HDL cholesterol through HDL receptor, SR-B1 (28).
Cholesterol enters the steroidogenic pathway by the action of the enzyme cholesterol esterase and transferred from the outer mitochondrial membrane of steroidogenic cells to the inner mitochondrial membrane by the steroid acute regulatory (STAR) protein. This transport is followed by the conversion of cholesterol to pregnenolone which is the first step of steroid synthesis and the major action of ACTH on adrenals (Figure 3). The conversion of cholesterol to pregnenolone requires the action of the cholesterol side-chain cleavage enzyme, commonly referred to as P450scc, encoded by the CYP11A gene located on chromosome 15 in mitochondria. This cleavage gives birth to 21-carbon (C21) molecules resulting from the C27 cholesterol molecule. These reactions require molecular oxygen and a pair of electrons. The electrons are donated by nicotinamide adenine dinucleotide phosphate (NADPH) to adrenodoxin reductase (flavoprotein) and then to adrenodoxin (an iron-sulfur protein) and finally to P450scc. Electron transport to microsomal cytochrome P450 involves the enzyme P450 reductase (Figure 4). The steroid hormones produced by the adrenal cortex are members of a large family of compounds derived from the cyclopentanoperhydrophenanthrene ring structure that comprises three cyclohexane rings and one cyclopentane ring. P450scc is needed for the production of all steroids in the human body, including those produced by the adrenal. It is expressed in all adrenal cortex zones, and although its expression is obligatory for the synthesis of C19 steroids (DHEA, DHEA-S, and A4) it is the presence of downstream enzymes that determines whether these cells produce C21 corticosteroids or C19 steroids (Figure 5).

Figure 3:
The role of the mitochondrion in the adrenal steroidogenesis.

Figure 4:
Reaction mechanism of hydroxylations catalyzed by cytochrome P-450s of adrenal cortex mitochondria. Abbreviations: XH = substrate; XOH = product; Fp = flavoprotein; ISp = iron-sulfur protein.

Figure 5:
Adrenal androgen biosynthetic pathway.
Biosynthesis Of Adrenal Androgens.
The second step of adrenal steroidogenesis is mediated by cytochrome P450 17A1 enzyme which acts both as a hydroxylase hydroxylating pregnenolone, and as a lyase splitting the C17–C20 bond of 17-hydroxypregnenolone (17OHP5), resulting in the production of DHEA (29-31). Specifically, CYP17A1 gene encodes a protein that catalyzes two metabolic pathways, the 17a-hydroxylation (principal for the androgen and glucocorticoid pathway) and the 17,20 lyase reaction (specific for androgen pathway). Even though the affinity of the human Cytochrome P450 17A1 is similar either for Δ5 steroid substrate (pregnenolone) or Δ4 steroid substrate (progesterone), the predominant pathway for the 17,20 lyase reaction is via the 17 OH pregnenolone (Δ5 substrate) (32). This 17,20-lyase activity is predominant in zona reticularis. There, the presence of cytochrome b5, form A enzyme (encoded by the CYB5A gene), a cofactor/regulator of cytochrome P450 17A1 function, promotes 17,20-lyase activity (33).
DHEA is then converted to DHEAS sulfate by an adrenal sulfokinase (encoded by the SULT2A1 gene). This enzyme, present mostly in the cytoplasm of adrenocortical cells in zona reticularis, mediates the sulfo conjunction of the Δ5 steroids (pregnenolone, 17α-hydroxypregnenolone, DHEA, and A5. Although all of these adrenal steroids, in fetal life, act as substrates for the corresponding sulfated products; in adult life the main substrate for the production of DHEAS is DHEA (34). During embryonic development DHEAS is supplied in maternal circulation from the fetal adrenals and acts as a substrate for estrogen synthesis from the placenta in such a way that the concentration of maternal estriol (produced in the placenta) reflects fetoplacental steroidogenesis (5). After birth however the sulfation of DHEA to DHEAS has a preventive role for androgen production by preventing excessive amounts of DHEA, a substrate for HSD3B2, to produce increased A4 and finally T (35). Of note, the expression of SULT2A1 in zona reticularis increases during adrenarche (36).
DHEA is also converted to A4 by the enzyme 3-β-hydroxysteroid dehydrogenase (3βHSD) encoded by the HSD3B2 gene. This pathway represents the predominant pathway for the production of DHEA in humans. 3-β-Hydroxysteroid dehydrogenase has a major role in the synthesis of androgens but also of mineralocorticoids and glucocorticoids as it catalyzes the conversion of Δ5 (pregnenolone, 17α-hydroxypregnenolone, DHEA and A5) to Δ4 steroids (progesterone, 17α-hydroxyprogesterone, Α4 and T). In fetal adrenal the expression of 3βHSD peaks at the 8th to 9th gestational week, resulting in the production of cortisol at 8th to 10th gestational week, decreasing thereafter and being undetectable at 14th gestational week. The decrease of HSD3B2 expression is followed by a decrease of cortisol synthesis. The transient cortisol synthesis by the 10th gestational week may exert a negative feedback on ACTH secretion suppressing adrenal synthesis of androgenic C19 steroids during the time of genital differentiation. Thus, in humans, early cortisol biosynthesis provides a mechanism to safeguard female sexual development (9,37). Given that fetal adrenal cell has a low expression of HSD3B2 gene until the end of 2nd trimester (38-39) it becomes evident that the fetal adrenal gland is more likely to produce Δ5 steroids, especially DHEA, than Δ4 steroids with mineralocorticoid and glucocorticoid activities. This low HSD3B2 expression is apparent also in adrenarche where the characteristic expansion of zona reticularis, demonstrating lower concentration of HSD3B2 compared to the adjacent zona fasciculata, facilitates the increased amount of DHEAS which marks the prepubertal to adult life transition (40-42,36).
Finally, Δ4 can be converted to T, although adrenal secretion of this hormone is minimal (Figure 5) (43-45). Human type 5 17β-hydroxysteroid dehydrogenase (encoded by the 17β-HSD) catalyzes the conversion of A4 to T (46-47). The fetal as well as the postnatal adrenal also expresses AKR1C3 gene in zona reticularis, which appears to be responsible for the small amount of T produced directly by the adrenal glands (48) and is likely responsible for the larger amounts of androgens produced in congenital adrenal hyperplasia.
CIRCULATION AND METABOLISM
Adrenal androgens are secreted from the adrenal cortex in an unbound state. Bound steroids are biologically inactive. Androstenedione, DHEA and DHEAS bind mainly to albumin. About 90% of adrenal androgens are bound to albumin and 3% approximately are bound to sex hormone-binding globulin (SHBG). The binding globulins have high affinity and low capacity, whereas, albumin has low affinity and high capacity for steroids. Adrenal androgens can follow two different pathways after entering the circulation. Their metabolism turns either towards degradation and inactivation or towards peripheral conversion to their more potent derivatives T and dihydrotestosterone (DHT). Adrenal androgens and their metabolites are inactivated or degraded in various tissues, including liver and kidney (49). Major biochemical routes for inactivation and excretion are conjugation of androgens to glucuronate or sulfate residues to produce hydrophilic glucuronides or sulfates, respectively, excreted in the urine (Figure 6A).
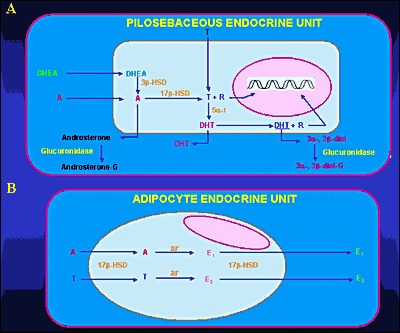
Figure 6:
Metabolism of adrenal androgens in the pilosebaceous unit (A) and adipocyte tissue (B). Abbreviations: A, Δ4-androstenedione; T, testosterone; DHT, dihydrotestosterone; R, receptor; 3β-HSD, 3β-hydroxysteroid dehydrogenase; 17β-HSD, 17β-hydroxysteroid dehydrogenase; 5α-r, 5α-reductase; 3α-, 3β -diol, 3α-androstenediol, 3β-androstenediol; 3α-, 3β- diol-G, 3α-diol-glucuronide, 3β-diol-glucuronide; androsterone-G, androsterone glucuronide; ar, aromatase; E1, estrone; E2, 17β-estradiol
DHEA, DHEAS, and A4 are converted to the potent androgens T and DHT in peripheral tissues. Major conversions are those of A4 to T and T to DHT by the enzymes 17β-hydroxysteroid dehydrogenase (17β-HSD) and 5α-reductase, respectively. Major peripheral sites of androgen conversion are the hair follicles, the sebaceous glands (Figure 6A), the prostate, the external genitalia and the adipose tissue (50-51). DHEAS is the sulfated version of DHEA. This conversion is catalyzed by sulfotransferase (SULT2A1) primarily in the adrenals, the liver, the kidney and small intestine. The concentrations of DHEAS in the circulation are about 300 times greater than those of free DHEA. The former show no diurnal variation, whereas the latter reach their peak in the early morning hours. DHEA secreted by the adrenal gland can be also converted to A4. Both DHEA and DHEAS are also metabolized to 7α and 16α – hydroxylated derivatives and by 17β reduction to Α5 and its sulfate. Androstenedione is converted either to T or by reduction of its 4,5-double bond to etiocholanolone or androsterone. Testosterone is converted to DHT in androgen-sensitive tissues by 5β reduction. The product is mainly metabolized by 3α reduction to 3α androstanediol. The metabolites of these androgens are conjugated either as glucuronides or sulfates and excreted in the urine. Active uptake of androgens and in situ estrogen synthesis occur in peripheral adipose tissue (Figure 6B) through the enzymes 17β-HSD and aromatase, respectively (52-55). Peripheral conversion contributes significantly to circulating T concentration in women, but not in men, in whom T is largely produced by the testis. Three main enzyme complexes are involved in the synthesis of estrogens in peripheral tissues (56-58):
- Aromatase for the aromatization of androstenedione to estrone.
- Estrone sulfatase (E1-STS), which catalyses the formation of estrone from estrone sulfate.
- Estradiol-17-β-hydroxysteroid dexydrogenase (17β-HSD) Type 1 which is responsible for the reduction of estrone to the biologically active estrogen, estradiol.
Finally, according to recent studies, 11-KT has been found to be the principal androgen made by the human adrenal. Both A4 and T may undergo 11-hydroxylation catalyzed by P450c11b (CYP11B1 gene) to yield 11OHA4 and 11OH-testosterone (11OH-T), respectively. These 11-hydroxysteroids may be oxidized by 11β-hydroxysteroid dehydrogenase type 2 (HSD-11B2), which is more known for its role in the oxidation of cortisol to cortisone, to 11-ketoandrostenedione (11-KA4) and 11-KT, respectively (Figure 7). These 11-keto steroids may then be 5α-reduced by 5α-reductase type 2 (SRD5A2 gene) in peripheral tissues, and possibly also by 5α-reductase type 1 (SRD5A1 gene) in the adrenal itself, to 5α-androstanedione and 5α-dihydrotestosterone (5αDHT), respectively. Both 11-KT and 11-ketodihydrotestosterone (11-KDHT) are bona fide androgens that bind to and transactivate the androgen receptor. Whereas most studies have addressed the synthesis of these steroids in castration resistant prostate cancer, other studies showed that they may have an important role also in other disease states (e.g. congenital adrenal hyperplasia).

Figure 7:
Novel Adrenal Androgens. 3bHSD2, 3b-hydroxysteroid dehydrogenase type 2; 11KA4, 11-ketoandrostenedione; 11KT, 11-ketotestosterone; 11OHA4, 11b-hydroxyandrostenedione; 11OHT, 11b-hydroxytestosterone; 17OH-PREG, 17a-hydroxypregnenolone; 17OH-PROG, 17a-hydroxyprogesterone; A4, androstenedione; AKR1C3, aldo-keto reductase 1C3; CYB5A, cytochrome b5; CYP11A1, cytochrome P450 cholesterol side-chain cleavage; CYP11B1, cytochrome P450 11b-hydroxylase; CYP17A1, cytochrome P450 17a-hydroxylase/17,-20-lyase; DHEA, dehydroepiandrosterone; DHEAS, DHEA sulfate; PREG, pregnenolone; StAR, steroidogenic acute regulatory protein; T, testosterone, SULT2A1, Sulfotransferase Family 2A Member 1; PROG, progesterone; CYP17A2 cytochrome P450 family 17 polypeptide 2; 11βHSD2, 11-β-hydroxysteroid dehydrogenase type 2.
ANDROGEN RECEPTOR
The inactive androgen precursors secreted by the adrenal glands are converted to T and DHT and exert their effects in most peripheral tissues by interacting with high-affinity receptor proteins. The androgen receptor (AR), member of the steroid receptor superfamily, also known as NR3C4 (nuclear receptor subfamily 3, group C, member 4) is a type of nuclear receptor that is activated by binding to T or DHT in the cytoplasm and then trans-locates into the nucleus. The AR is most closely related to the progesterone receptor, while progestins in higher dosages can block AR. Androgen receptors are encoded by the AR gene located on the X-chromosome at Xq11-12 (59). This gene contains a polymorphic CAG microsatellite repeat within exon 1, encoding for a variable length of polyglutamine chain at the amino terminal, the transactivation domain of the AR protein. Triplet-repeat DNA sequences can be sites of genetic instability, and their expansion in a variety of genes has been associated with human genetic diseases, such as fragile X-syndrome (60-61) and myotonic dystrophy (62). In the case of the AR gene, an inverse correlation of the number of CAG repeats with the risk for prostate cancer was described (63-66) and its expansion was documented in Kennedy's disease (spinal and bulbar muscular atrophy), a disorder associated with primary hypogonadism due to androgen insensitivity (67). In vitro studies showed that progressive expansion of the repeat length in the AR was associated with a linear decrease in its transactivation function (68). These observations support the idea that there is an optimal number of repeats, which varies in the population from 11 to 31 (average size: 21±2) (63). Methylation of deoxycytosine residues is another process involved in the modulation of gene expression. Belmont et al. (69) showed that the methylation of HpaII and HhaI sites near the polymorphic CAG repeats in the first exon of the human AR (HUMARA) locus correlated with X-inactivation.
Patients with idiopathic hirsutism were shown to have a normal number of CAG repeats but with a preponderance of the shortest and most active alleles (70). These patients had also a preferential methylation of the longer AR allele compared to normal subjects, leading to inactivation of the functionally weaker gene. This skewing could allow the shorter, more active AR allele (64,68) to be preferentially expressed explaining the peripheral hypersensitivity to androgens in hirsute patients.
Multiple "coactivators" were identified enhancing transcription of the AR gene (71) including AP-1 (72), Smad3 (73-74), nuclear factor kB (NF-kB) (75-76) sex-determining region Y (SRY) (77) and the Ets family of transcription factors (78). The relative importance of these molecules for any particular cell type remains unclear, since the ability of a putative coregulator to alter the transcriptional activity is typically examined in transient transfection experiments. Although AR is normally thought to function as a homodimer, it was also shown to heterodimerize with other nuclear receptors including the estrogen receptor (ER) (79) glucocorticoid receptor (GR) (80) and testicular orphan receptor 4 (TR4) (81). One of the major mechanisms through which coregulators might function is by forming a bridge between the DNA-bound nuclear receptor and the basal transcriptional machinery (type I regulators) (82). Coactivators may also facilitate ligand binding, promote receptor nuclear translocation, or mediate signal transduction (type II coregulators). The role of "corepressors" in AR function is poorly defined. Three corepressors of androgen-bound AR have been identified to date, cyclin D1, calreticulin, and HBO1. However, relatively little is known about the mechanism of their repressive effect.
ADRENAL ANDROGEN PHYSIOLOGY AND REGULATION
Regulation
Adrenal androgens are secreted by the adrenal glands in response to ACTH, a 39-amino acid peptide synthesized and secreted by the anterior pituitary (Figure 8). It is derived from proopiomelanocortin (POMC), a large precursor molecule from which β-lipotropin hormone and β-endorphin are also derived (83-84). ACTH is the predominant form of corticotropin in plasma and has a half-life of approximately 10 minutes (85). Its synthesis and secretion are primarily regulated by corticotropin-releasing hormone (CRH) and arginine-vasopressin (AVP), both of which are produced by parvocellular neurons of the paraventricular nucleus of the hypothalamus and act in synergy with each other (86-87). Under ACTH regulation, adrenal androgens are secreted synchronously with cortisol. There are three mechanisms of neuroendocrine control: [1] episodic secretion and the circadian rhythm of ACTH, [2] stress responsiveness of the hypothalamic-pituitary-adrenal axis (HPA), [3] feedback inhibition of ACTH secretion by cortisol. [1]

Figure 8:
Schematic presentation of the adrenal androgen regulation. Downloaded from: wikis.lib.ncsu.edu
The circadian rhythm is the result of the central nervous system regulation of CRH and ACTH nyctohemeral secretory episodes. The major secretory episodes begin in the sixth to eighth hour of sleep and then begin to decline as wakefulness occurs. Cortisol secretion then gradually declines during the day with fewer secretory episodes (88). The circadian rhythm of adrenal androgens is typical in different physiologic and pathologic conditions. Patients with nonclassical 21-hydroxylase deficiency, for example, have a distinct pattern of adrenal steroid secretion characterized by a high-frequency 17-hydroxyprogesterone release accompanied by a relative nocturnal cortisol deficiency (89-90). [2] Plasma ACTH and cortisol secretion are secreted within minutes following the onset of physical stress. This response abolishes circadian periodicity if the stress is prolonged. Stress responses originate in the CNS and result to CRH and ACTH secretion. [3] Corticotropin-stimulated cortisol exerts major feedback inhibitory influences at the concentration of both the hypothalamus and the anterior pituitary by suppressing CRH and ACTH synthesis and secretion.
Plasma DHEA, A4, and T concentrations parallel closely the circadian rhythm of plasma cortisol. Plasma DHEA-S concentrations do not exhibit a circadian rhythm because of the much longer circulating half-life of this sulfated steroid (91-92). Numerous other endocrine signals (93) were proposed as coregulators of adrenal androgen secretion. Among these are prolactin (PRL) (94), estrogen (95-99), epidermal growth factor (100), prostaglandins (101), angiotensin (102), GH (103), gonadotropins (104-105), β-lipotropin, and β-endorphin. Glasow et al. reported the presence of PRL receptors in the human adrenal gland and suggested a direct effect of PRL on adrenal steroidogenesis that may be of particular relevance in clinical disorders characterized by hyperprolactinemia (106). Interestingly, adults with hyperprolactinemia have increased secretion of AAs by the zona reticularis, which is corrected by reduction of PRL secretion with bromocriptine (107). In women with PRL-secreting tumors there is a correlation between PRL concentration and DHEA-S (108). Pabon et al. (109) have detected the presence of LH- HCG receptors in zona reticularis and fasciculata. The receptor bearing cells were positive for steroidogenic enzymes, indicating that the receptors could be coupled to DHEAS secretion (110-112).
Cytokines interfere with steroidogenesis at the level of the adrenals, testes and ovaries. Within the adrenal adrenocortical and chromaffin cells cytokines such as interleukin (IL) 1, IL-6, tumor necrosis factor (TNF), leukemia inhibitory factor (LIF) and IL-18 are produced. They have a key role in the immune-adreno-cortical communication. Thus, in autoimmune and inflammatory diseases an adequate adrenal stress response is observed. In addition, cytokines such as IL-8 and monocyte chemotactic protein-1 (MCP-1) are involved in steroidogenesis (113). ΙL-6 also is known to activate the HPA axis by stimulating both the CRH and the AVP -secreting neurons of the paraventricular nucleus of the hypothalamus, and their terminals at the median eminence, the corticotrophs of the anterior pituitary, and the cortisol-secreting adrenal cells in rats. In the latter it acts through specific receptors expressed mainly in the zona fasciculata and reticularis, but also with lower density in the zona glomerulosa (114-115). The ability of IL-6 to stimulate glucocorticoids, mineralocorticoids, and androgens suggests that this cytokine might have a role in coordinating the response of all adrenocortical zones. Its secretion is regulated by different substances, such as CRH, ACTH, angiotensin II, or immune products such as IL-1 α/β indicating that IL-6 may play a major role in the interaction of the adrenal function with the immune/inflammatory reaction (116).
Interleukin 1 and TNF regulate the activity of HPA axis at several levels. Studies investigated their action on adrenal steroidogenesis and indicated that IL-1α and IL-1β increase cortisol, A4, DHEA, DHEAS production and the accumulation of mRNAs for STAR, 17α-hydroxylase/17,20-lyase (CYP17A1) and HSD3B2 in these cells. TNF induced cortisol production (117).
Both ACTH and PRL stimulate AAs secretion by the fetal adrenal zone. In addition, placental CRH appears to play a major role in sustaining this zone and stimulating androgen secretion together with corticotropin and/or PRL (118).
Physiology
Adrenal androgens are secreted in small amounts during infancy and early childhood. DHEAS is maintained at minimum concentrations for 5 years in both male and females, after which a gradual increase is observed (115). Their secretion gradually increases with age, paralleling the growth of zona fasciculata and zona reticularis. Disturbances in both enzymatic activity in zona fasciculata and zona reticularis and its regulators (ACTH or peptides of hypothalamic – pituitary origin, such as PRL) may result in syndromes of hirsutism and virilization in females. Adrenal cortex normally secretes androgens in increasing amounts beginning at about 6-7 years of age in girls and 7-8 years of age in boys. This rise continues until late puberty. Adrenarche (secretion of adrenal androgens) occurs years before gonadarche (secretion of gonadal sex steroids). The appearance of pubic hair (pubarche) results from a rise in adrenal androgen concentration (adrenarche) (116-117). The mechanism(s) by which zona fasciculata and zona reticularis develop with age, as well as the regulation of adrenarche onset are not understood. The biochemical hallmark of adrenarche is accelerated DHEAS production from the adrenal gland. The axillary and pubic hair regions are the most sensitive androgen-dependent regions and they represent the clinical manifestation of adrenarche. Children with premature pubarche demonstrate hormonal responses to CRH stimulation test similar in magnitude to those of prepubertal children of comparable age, ruling out a prominent role of CRH in premature pubarche (118). Gell et al. suggested that as children mature, a decrease of HSD3B2 activity in the adrenal zona reticularis occurs, leading to an increased production of DHEA and DHEA-S, as seen during adrenarche, by shifting pregnenolone through the 17α-hydroxylase/17, 20 lyase pathway (Figure 5) (39).
Activation of the type 1 insulin-like growth factor (IGF1) receptor was shown to enhance steroidogenic responsiveness of the fetal zone cells to ACTH by modulating the ACTH signal transduction pathway at some point downstream from ACTH receptor binding (119). Also, locally produced IGF2 modulates fetal adrenocortical cells function by increasing responsiveness to ACTH via activation of the IGF1 receptor and increases the capacity of those cells for androgen synthesis by directly augmenting the expression of P450c17 (119). Thus, IGF2 may play a pivotal role in AA production, both physiologically in utero and at adrenarche, as well as in conditions of hyperandrogenemia (119). All together, these data indicate that the IGF system is important in the regulation of the differential function of adult human adrenocortical cells (120). The rise in plasma concentrations of the AAs at adrenarche occurs in the presence of constant cortisol concentrations, suggesting that factors other than corticotropin are involved. The influences of sex and age are minor in the modulation of adrenal steroidogenesis supporting the concept that extra-adrenal factors prevail in the differential modulation of AAs and cortisol (121). These may include POMC-derived or other still uncovered peptides. An increased serine phosphorylation of human P450c17 might have a role in the development of both the excessive adrenarche and hyperandrogenism of patients with the polycystic ovary syndrome (PCOS) resulting in a substantial increase in 17-20-lyase activity (122-124) (Figure 5). P450c17 is the key enzyme that regulates androgen synthesis. (125). It is the only enzyme known to be able to convert C21-precursors to the androgen pro-hormones, the 17-ketosteroids. It is a single enzyme with two activities, 17 a-hydroxylase and 17,20-lyase (Figure 5) and serine phosphorylation appears to modulate the activity of P450c17. In particular, it promotes the 17,20-lyase activity, and at the same time it inhibits the activity of the insulin receptor (123-124,126-128). It was postulated that a single abnormal serine kinase might hyperphosphorylate both P450c17 and the insulin receptor, accounting for the hyperandrogenism and hyperinsulinism responsible for both premature pubarche and PCOS later in life (129). In vitro studies, however, failed to find evidence for increased autophosphorylation of the insulin receptor-β subunit and P450c17 in PCOS (130). The reason for this might be related to the many different factors needed for P450c17 optimal activity and not normally expressed in the cell line used for that study (48).
BIOLOGIC EFFECTS
In adult men, the conversion of adrenal A4 to T accounts for less than 5% of the production rate of the latter, making its participation in the physiologic androgenization of the male negligible. Excessive AA secretion appears to have no major clinical consequences in the adult man, although this may be debated. Adrenal androgens hypersecretion in prepubertal boys, on the other hand has clearly been associated with isosexual precocious puberty.
In adult women, adrenal A4 and A4 generated from peripheral conversion of DHEA contribute substantially to total androgen production and effects. In the follicular phase of the menstrual cycle, adrenal precursors account for two thirds of T production and half of DHT production. At midcycle, the ovarian contribution increases, and the adrenal precursors account for 40% of T production. In women, increased AA production may be manifested as cystic acne, hirsutism, male type baldness, menstrual irregularities, oligoovulation or anovulation, infertility, and/or frank virilization. Excessive adrenal androgen secretion in prepubertal or pubertal girls can cause heterosexual precocious puberty.
Abnormalities in the timing and intensity of adrenarche are associated with PCOS, congenital adrenal hyperplasia (CAH) and insulin resistance conditions. Recently, we have shown that in postmenopausal PCOS women, androgen concentration at baseline are greater in PCOS than control women and remain increased after ACTH stimulation, while the results of the dexamethasone suppression test in postmenopausal PCOS women suggest that DHEAS and total T are partially of adrenal origin (131). Although the ovarian contribution was not fully assessed, increased A4 production suggests that the ovary also contributes to hyperandrogenism in postmenopausal PCOS women. In conclusion, this study indicates that postmenopausal PCOS women are exposed to higher adrenal and ovarian androgen concentrations than non-PCOS women (131).
Studies conducted over the past few years have investigated the use of DHEA to treat female infertility (132-133). Women with poor ovarian reserve, after DHEA supplementation 4 to 12 weeks prior an in vitro fertilization (IVF) cycle, had a 50-80% reduction in miscarriages (134). However, its efficacy in treating infertility remains controversial (135-137).
Reports demonstrate DHEA as a replacement therapy in the elderly (138-139). At 70-80 years of age, peak DHEA concentrations are about 10-20% of those in young adults. These reports suggest DHEA as replacement treatment in menopausal women; it has been reported to restore both the androgenic and estrogenic environment and reduce most of the symptoms of menopause (140-142). Other reports have suggested that oral DHEA in doses of 25-50 mg/d may restore plasma T concentrations to normal in some women with hypopituitarism who have diminished libido despite adequate estrogen therapy (143-145). In addition, DHEA replacement therapy has been investigated for the conditions of adrenopause and adrenal insufficiency (146-149). In spite of these few reports so far DHEA does not appear to be effective for perimenopausal symptoms (135) nor has it been shown to be effective as an “anti-aging” agent, as its effects in trials on cognitive function, body composition, insulin resistance, and well-being have been inconsistent (150-157,146). Based upon available data, the Endocrine Society guidelines, suggested against the routine use of DHEA for sexual function (or other indications) in postmenopausal women because of its limited efficacy and lack of long-term safety data (158). Clinical trial data on the efficacy of DHEA therapy in women with primary adrenal insufficiency are mixed.
In several studies of women with premature ovarian insufficiency (POI), serum ovarian androgen concentrations (A4 and/or T) were lower than those of age-matched women without ovarian insufficiency, but similar to those seen in older postmenopausal women (159-161). In contrast, DHEAS concentrations were normal (although they would be expected to be low in those women with coexisting primary adrenal insufficiency). Potential side effects of androgen replacement include hirsutism and acne, and with oral preparations (e.g. DHEA), dyslipidemia. However, in women with autoimmune ovarian failure and coexisting adrenal insufficiency, adrenal androgen therapy with DHEA may be beneficial.
Other studies investigate the role of DHEA and DHEAS in the immune response and suggest that adrenal androgens have opposite biological effects to those of corticosteroids (162). DHEA is a cortisol antagonist (163). Research studies indicate DHEA supplementation has an anti-depressant effect (164-166). Exogenous DHEA has been proposed to have a number of potential benefits (on sexual function, depression, cognition, and inflammation), but available clinical trial data do not support these claims (115,167-168,137). It is widely available in some countries as a dietary supplement; however, quality control of these products has been shown to be quite poor (169,170).
Both gonadal and AAs contribute to the positive impact of androgenic steroids on bone cell metabolism in vitro (171). Interestingly, a study found that the potential anabolic effect of androgens on bone might not be mediated at the level of the mature osteoblast but at the level of fetal, less differentiated, osteoblastic cell lines (172).
Finally, although A4 seems to increase serum T and estrone concentrations when administered acutely to women, (173) the impact of regular use on sexual function or its potential androgenic side effects in women are unknown
CONCLUSIONS
The physiology of adrenal androgens follows the different periods of life starting from the fetal period. During this period, the secretion of these hormones from the fetal adrenal is important. It is not clarified as yet its role in the fetal development or survival, while it is of major importance for parturition. DHEA is the most prevalent steroid hormone in the body. After birth DHEA(S) concentrations fall rapidly with the involution of the fetal adrenal and rise slowly during childhood accelerating at adrenarche before the onset of puberty. The physiology of adrenarche is well described although its trigger has not been identified yet. DHEA concentrations drop dramatically with aging. There are pronounced differences in the average DHEA concentrations between men and women, with women on average having lower DHEA concentrations. The spectrum of women and men that would benefit from DHEA therapy is not clearly defined. Further studies are needed to investigate the side effects of the DHEA replacement therapy and to define the range of dosage that is more effective without complications. During menopause transition mean circulating DHEAS concentrations exhibit a positive inflection starting in the early perimenopause, continuing through the early post menopause and returning to early perimenopausal concentrations by late post menopause. This rise in mean DHEAS is accompanied by concomitant rises in T, DHEA, A4, and an equal rise in A5. Studies have shown that the mean A4 and T concentrations changed the least while mean DHEAS and A5 changed the most. The role of these changes in altering the estrogen/androgen balance in menopause is not known.
REFERENCES
- 1.
- Rege J, Nakamura Y, Satoh F, et al. Liquid chromatography-tandem mass spectrometry analysis of human adrenal vein 19-carbon steroids before and after ACTH stimulation. The Journal of clinical endocrinology and metabolism. 2013;98:1182–1188. [PMC free article: PMC3590473] [PubMed: 23386646]
- 2.
- Kaufman FR, Stanczyk FZ, Matteri RK, et al. Dehydroepiandrosterone and dehydroepiandrosterone sulfate metabolism in human genital skin. Fertility and sterility. 1990;54:251–254. [PubMed: 2143146]
- 3.
- Luu-The V. Assessment of steroidogenesis and steroidogenic enzyme functions. J Steroid Biochem Mol Biol. 2013;137:176–182. [PubMed: 23770321]
- 4.
- Pelletier G. Expression of steroidogenic enzymes and sex-steroid receptors in human prostate. Best practice & research Clinical endocrinology & metabolism. 2008;22:223–228. [PubMed: 18471781]
- 5.
- Rainey WE, Rehman KS, Carr BR. The human fetal adrenal: making adrenal androgens for placental estrogens. Seminars in reproductive medicine. 2004;22:327–336. [PubMed: 15635500]
- 6.
- Rosenfield RL. Hirsutism and the variable response of the pilosebaceous unit to androgen. The journal of investigative dermatology Symposium proceedings/the Society for Investigative Dermatology, Inc [and] European Society for Dermatological Research. 2005; 10:205–208. [PubMed: 16382665]
- 7.
- Longcope C. Adrenal and gonadal androgen secretion in normal females. In: Horton R, Lobo RA, eds. Clinics in Endocrinology and Metabolism. Philadelphia: W.B. Saunders, 1986:213-228. [PubMed: 3013468]
- 8.
- Hanley NA, Rainey WE, Wilson DI, et al. Expression profiles of SF-1, DAX1, and CYP17 in the human fetal adrenal gland: potential interactions in gene regulation. Mol Endocrinol. 2001;15:57–68. [PubMed: 11145739]
- 9.
- Goto M, Piper Hanley K, Marcos J, et al. In humans, early cortisol biosynthesis provides a mechanism to safeguard female sexual development. J Clin Invest. 2006;116:953–960. [PMC free article: PMC1421344] [PubMed: 16585961]
- 10.
- Mesiano S, Coulter CL, Jaffe RB. Localization of cytochrome P450 cholesterol side-chain cleavage, cytochrome P450 17 α-hydroxylase/17, 20-lyase, and 3β-hydroxysteroid dehydrogenase isomerase steroidogenic enzymes in human and rhesus monkey fetal adrenal glands: reappraisal of functional zonation. J Clin Endocrinol Metab. 1993;77:1184–1189. [PubMed: 8077311]
- 11.
- McNutt NS, Jones AL. Observations on the ultrastructure of cytodifferentiation in the human fetal adrenal cortex. Lab Invest. 1970;22:513–527. [PubMed: 5423978]
- 12.
- Sucheston ME, Cannon MS. Development of zonular patterns in the human adrenal gland. J Morphol. 1968;126:477–491. [PubMed: 5716437]
- 13.
- Keene MF, Hewer EE. Observations on the development of the human suprarenal gland. J Anat. 1927;61:302–324. [PMC free article: PMC1249958] [PubMed: 17104143]
- 14.
- Grueters A, Korth-Schutz S. Longitudinal study of plasma dehydroepiandrosterone sulfate in preterm and fullterm infants. J Clin Endocrinol Metab. 1982;55:314–320. [PubMed: 6211463]
- 15.
- Honour JH, Wickramaratne K, Valman HB. Adrenal function in preterm infants. Biol Neonate. 1992;61:214–221. [PubMed: 1610949]
- 16.
- Kojima S, Yanaihara T, Nakayama T. Serum steroid levels in children at birth and in early neonatal period. Am J Obstet Gynecol. 1981;140:961–965. [PubMed: 6455919]
- 17.
- Wiener D, Smith J, Dahlem S, et al. Serum adrenal steroid levels in healthy full-term 3-day-old infants. J Pediatr. 1987;110:122–124. [PubMed: 3794871]
- 18.
- Lanman JT. The fetal zone of the adrenal gland: its developmental course, comparative anatomy, and possible physiologic functions. Medicine (Baltimore). 1953;32:389–430. [PubMed: 13110603]
- 19.
- Bocian-Sobkowska J. Morphometric study of the human suprarenal gland in the first postnatal year. Folia Morphol (Warsz). 2000;58:275–284. [PubMed: 11000884]
- 20.
- Walter L. Miller. Steroidogenesis: Unanswered Questions Review. Trends Endocrinol Metab. 2017 Nov;28(11):771–793. [PubMed: 29031608]
- 21.
- Parker CRJ, Mixon RL, Brissie RM, et al. Aging alters zonation in the adrenal cortex of men. J.Clin.Endocrinol.Metab. 1997;82:3898–3901. [PubMed: 9360559]
- 22.
- Auchus RJ, Rainey WE. Adrenarche - physiology, biochemistry and human disease. Clin Endocrinol (Oxf). 2004 Mar;60(3):288–96. Review. [PubMed: 15008992]
- 23.
- Auchus RJ. The physiology and biochemistry of adrenarche. Endocr Dev. 2011;20:20–7. [PubMed: 21164255]
- 24.
- Guyton Physiology 11th ed. Chapter 77. Adrenocortical Hormones.
- 25.
- Davis JO. Regulation of aldosterone secretion. In: Einstein AB e, ed. The adrenal cortex. Boston: Little, Brown, 1967:203-247.
- 26.
- Labrie F, Belanger A, Cusan L, et al. Marked decline in serum concentrations of adrenal C19 sex steroid precursors and conjugated androgen metabolites during aging. J. Clin.Endocrinol. Metab. 1997;82:2396–2402. [PubMed: 9253307]
- 27.
- Kronenberg. Williams Textbook of Endocrinology 11th ed. Chapter 14. The adrenal cortex.
- 28.
- Hoekstra M., Van Berkel Theo JC., Van Eck M. A multi-purpose player in cholesterol and steroid metabolism. World J Gastroenterol. 2010 December 21;16(47):5916–5924. [PMC free article: PMC3007109] [PubMed: 21157967]
- 29.
- Bradshaw KD, Waterman MR, Couch RT, et al. Characterization of complementary deoxyribonucleic acid for human adrenocortical 17 alpha-hydroxylase: a probe for analysis of 17 alpha- hydroxylase deficiency. Mol Endocrinol. 1987;1(5):348–354. [PubMed: 3274893]
- 30.
- Kagimoto M, Winter JS, Kagimoto K, et al. Structural characterization of normal and mutant human steroid 17 alpha- hydroxylase genes: molecular basis of one example of combined 17 alpha- hydroxylase/17,20 lyase deficiency. Mol Endocrinol. 1998;2(6):564–570. [PubMed: 2843762]
- 31.
- Swart P, Estabrook RW, Mason JI, et al. Catalytic activity of human and bovine adrenal cytochromes P-450 17 alpha, lyase expressed in Cos 1 cells. Biochem Soc Trans. 1989;17(6):1025–1026. [PubMed: 2628057]
- 32.
- Auchus RJ, Lee TC, Miller WL. Cytochrome b5 augments the 17,20-lyase activity of human P450c17 without direct electron transfer. J Biol Chem. 1998;273:3158–3165. [PubMed: 9452426]
- 33.
- Turcu, et al. Adrenal androgens and androgen precursors: definition, synthesis, regulation and physiologic actions. Compr Physiol. 2014 Oct;4(4):1369–1381. [PMC free article: PMC4437668] [PubMed: 25428847]
- 34.
- Rainey WE, Nakamura Y. Regulation of the adrenal androgen biosynthesis. The Journal. [PMC free article: PMC2699571] [PubMed: 17945481]
- Adrenal Androgens - EndotextAdrenal Androgens - Endotext
Your browsing activity is empty.
Activity recording is turned off.
See more...