The routes that lead inward from the cell surface to lysosomes start with the process of endocytosis, by which cells take up macromolecules, particulate substances, and, in specialized cases, even other cells. In this process, the material to be ingested is progressively enclosed by a small portion of the plasma membrane, which first invaginates and then pinches off to form an endocytic vesicle containing the ingested substance or particle. Two main types of endocytosis are distinguished on the basis of the size of the endocytic vesicles formed. One type is called phagocytosis (“cellular eating”), which involves the ingestion of large particles, such as microorganisms or dead cells via large vesicles called phagosomes (generally >250 nm in diameter). The other type is pinocytosis (“cellular drinking”), which involves the ingestion of fluid and solutes via small pinocytic vesicles (about 100 nm in diameter). Most eucaryotic cells are continually ingesting fluid and solutes by pinocytosis; large particles are most efficiently ingested by specialized phagocytic cells.

Specialized Phagocytic Cells Can Ingest Large Particles
Phagocytosis is a special form of endocytosis in which large particles such as microorganisms and dead cells are ingested via large endocytic vesicles called phagosomes. In protozoa, phagocytosis is a form of feeding: large particles taken up into phagosomes end up in lysosomes, and the products of the subsequent digestive processes pass into the cytosol to be utilized as food. However, few cells in multicellular organisms are able to ingest such large particles efficiently. In the gut of animals, for example, the particles of food are broken down extracellularly and their hydrolysis products are imported into cells.
Phagocytosis is important in most animals for purposes other than nutrition, and it is mainly carried out by specialized cells—so-called professional phagocytes. In mammals, three classes of white blood cells act as professional phagocytes—macrophages, neutrophils, and dendritic cells. These cells all develop from hemopoietic stem cells (discussed in Chapter 22), and they defend us against infection by ingesting invading microorganisms. Macrophages also have an important role in scavenging senescent cells and cells that have died by apoptosis (discussed in Chapter 17). In quantitative terms, the latter function is by far the most important: our macrophages phagocytose more than 1011 senescent red blood cells in each of us every day, for example.
Whereas the endocytic vesicles involved in pinocytosis are small and uniform, phagosomes have diameters that are determined by the size of the ingested particle, and they can be almost as large as the phagocytic cell itself (). The phagosomes fuse with lysosomes inside the cell, and the ingested material is then degraded. Any indigestible substances will remain in lysosomes, forming residual bodies. Some of the internalized plasma membrane components never reach the lysosome, because they are retrieved from the phagosome in transport vesicles and returned to the plasma membrane.
Phagocytosis by a macrophage. A scanning electron micrograph of a mouse macrophage phagocytosing two chemically altered red blood cells. The red arrows point to edges of thin processes (pseudopods) of the macrophage that are extending as collars to engulf (more...)
To be phagocytosed, particles must first bind to the surface of the phagocyte. However, not all particles that bind are ingested. Phagocytes have a variety of specialized surface receptors that are functionally linked to the phagocytic machinery of the cell. Unlike pinocytosis, which is a constitutive process that occurs continuously, phagocytosis is a triggered process, requiring that receptors be activated that transmit signals to the cell interior and initiate the response. The best-characterized triggers are antibodies, which protect us by binding to the surface of infectious microorganisms to form a coat in which the tail region of each antibody molecule, called the Fc region, is exposed on the exterior (discussed in Chapter 24). This antibody coat is recognized by specific Fc receptors on the surface of macrophages and neutrophils, whose binding induces the phagocytic cell to extend pseudopods that engulf the particle and fuse at their tips to form a phagosome ().
Phagocytosis by a neutrophil. An electron micrograph of a neutrophil phagocytosing a bacterium, which is in the process of dividing. (Courtesy of Dorothy F. Bainton, Phagocytic Mechanisms in Health and Disease. New York: Intercontinental Book Corporation, (more...)
Several other classes of receptors that promote phagocytosis have been characterized. Some recognize complement components, which collaborate with antibodies in targeting microbes for destruction (discussed in Chapter 25). Others directly recognize oligosaccharides on the surface of certain microorganisms. Still others recognize cells that have died by apoptosis. Apoptotic cells lose the asymmetric distribution of phospholipids in their plasma membrane. As a consequence, negatively charged phosphatidylserine, which is normally confined to the cytosolic leaflet of the lipid bilayer, is now exposed on the outside of the cell, where it triggers the phagocytosis of the dead cell.
Remarkably, macrophages will also phagocytose a variety of inanimate particles—such as glass, latex beads, or asbestos fibers—yet they do not phagocytose live animal cells. It seems that living animal cells display “don't-eat-me” signals in the form of cell-surface proteins that bind to inhibiting receptors on the surface of macrophages. The inhibitory receptors recruit tyrosine phosphatases that antagonize the intracellular signaling events required to initiate phagocytosis, thereby locally inhibiting the phagocytic process. Thus phagocytosis, like many other cell processes, depends on a balance between positive signals that activate the process and negative signals that inhibit it.
Pinocytic Vesicles Form from Coated Pits in the Plasma Membrane
Virtually all eucaryotic cells continually ingest bits of their plasma membrane in the form of small pinocytic (endocytic) vesicles, which are later returned to the cell surface. The rate at which plasma membrane is internalized in this process of pinocytosis varies between cell types, but it is usually surprisingly large. A macrophage, for example, ingests 25% of its own volume of fluid each hour. This means that it must ingest 3% of its plasma membrane each minute, or 100% in about half an hour. Fibroblasts endocytose at a somewhat lower rate (1% per minute), whereas some amoebae ingest their plasma membrane even more rapidly. Since a cell's surface area and volume remain unchanged during this process, it is clear that the same amount of membrane that is being removed by endocytosis is being added to the cell surface by exocytosis, the converse process, as we discuss later. In this sense, endocytosis and exocytosis are linked processes that can be considered to constitute an endocytic-exocytic cycle.
The endocytic part of the cycle often begins at clathrin-coated pits. These specialized regions typically occupy about 2% of the total plasma membrane area. The lifetime of a clathrin-coated pit is short: within a minute or so of being formed, it invaginates into the cell and pinches off to form a clathrin-coated vesicle (). It has been estimated that about 2500 clathrin-coated vesicles leave the plasma membrane of a cultured fibroblast every minute. The coated vesicles are even more transient than the coated pits: within seconds of being formed, they shed their coat and are able to fuse with early endosomes. Since extracellular fluid is trapped in clathrin-coated pits as they invaginate to form coated vesicles, any substance dissolved in the extracellular fluid is internalized—a process called fluid-phase endocytosis.
The formation of clathrin-coated vesicles from the plasma membrane. These electron micrographs illustrate the probable sequence of events in the formation of a clathrin-coated vesicle from a clathrin-coated pit. The clathrin-coated pits and vesicles shown (more...)
Not All Pinocytic Vesicles Are Clathrin-coated
In addition to clathrin-coated pits and vesicles, there are other, less well-understood mechanisms by which cells can form pinocytic vesicles. One of these pathways initiates at caveolae (from the Latin for “little cavities”), originally recognized by their ability to transport molecules across endothelial cells, which form the inner lining of blood vessels. Caveolae are present in the plasma membrane of most cell types, and in some of these they are seen as deeply invaginated flasks in the electron microscope (). They are thought to form from lipid rafts, which are patches of the plasma membrane that are especially rich in cholesterol, glycosphingolipids, and GPI-anchored membrane proteins (see ). The major structural protein in caveolae is caveolin, a multipass integral membrane protein that is a member of a heterogeneous protein family.
Caveolae in the plasma membrane of a fibroblast. (A) This electron micrograph shows a plasma membrane with a very high density of caveolae. Note that no cytosolic coat is visible. (B) This rapid-freeze deep-etch image demonstrates the characteristic “cauliflower” (more...)
In contrast to clathrin-coated and COPI- or COPII-coated vesicles, caveolae are thought to invaginate and collect cargo proteins by virtue of the lipid composition of the calveolar membrane, rather than by the assembly of a cytosolic protein coat. Caveolae pinch off from the plasma membrane and can deliver their contents either to endosome-like compartments or (in a process called transcytosis, which is discussed later) to the plasma membrane on the opposite side of a polarized cell. Some animal viruses also enter cells in vesicles derived from caveolae. The viruses are first delivered to an endosome-like compartment, from where they are moved to the ER. In the ER, they extrude their genome into the cytosol to start their infectious cycle. It remains a mystery how material endocytosed in caveolae-derived vesicles can end up in so many different locations in the cell.
Cells Import Selected Extracellular Macromolecules by Receptor-mediated Endocytosis
In most animal cells, clathrin-coated pits and vesicles provide an efficient pathway for taking up specific macromolecules from the extracellular fluid. In this process, called receptor-mediated endocytosis, the macromolecules bind to complementary transmembrane receptor proteins, accumulate in coated pits, and then enter the cell as receptor-macromolecule complexes in clathrin-coated vesicles (see ). Receptor-mediated endocytosis provides a selective concentrating mechanism that increases the efficiency of internalization of particular ligands more than a hundredfold, so that even minor components of the extracellular fluid can be specifically taken up in large amounts without taking in a correspondingly large volume of extracellular fluid. A particularly well-understood and physiologically important example is the process whereby mammalian cells take up cholesterol.
Many animals cells take up cholesterol through receptor-mediated endocytosis and, in this way, acquire most of the cholesterol they require to make new membrane. If the uptake is blocked, cholesterol accumulates in the blood and can contribute to the formation in blood vessel walls of atherosclerotic plaques, deposits of lipid and fibrous tissue that can cause strokes and heart attacks by blocking blood flow. In fact, it was through a study of humans with a strong genetic predisposition for atherosclerosis that the mechanism of receptor-mediated endocytosis was first clearly revealed.
Most cholesterol is transported in the blood as cholesteryl esters in the form of lipid-protein particles known as low-density lipoproteins (LDL) (). When a cell needs cholesterol for membrane synthesis, it makes transmembrane receptor proteins for LDL and inserts them into its plasma membrane. Once in the plasma membrane, the LDL receptors diffuse until they associate with clathrin-coated pits that are in the process of forming (). Since coated pits constantly pinch off to form coated vesicles, any LDL particles bound to LDL receptors in the coated pits are rapidly internalized in coated vesicles. After shedding their clathrin coats, the vesicles deliver their contents to early endosomes, which are located near the cell periphery. Once the LDL and LDL receptors encounter the low pH in the endosomes, LDL is released from its receptor and is delivered via late endosomes to lysosomes. There the cholesteryl esters in the LDL particles are hydrolyzed to free cholesterol, which is now available to the cell for new membrane synthesis. If too much free cholesterol accumulates in a cell, the cell shuts off both its own cholesterol synthesis and the synthesis of LDL receptor proteins, so that it ceases either to make or to take up cholesterol.
A low-density lipoprotein (LDL) particle. Each spherical particle has a mass of 3 × 106 daltons. It contains a core of about 1500 cholesterol molecules esterified to long-chain fatty acids that is surrounded by a lipid monolayer composed of about (more...)
Normal and mutant LDL receptors. (A) LDL receptor proteins binding to a coated pit in the plasma membrane of a normal cell. The human LDL receptor is a single-pass transmembrane glycoprotein composed of about 840 amino acids, only 50 of which are on the (more...)
This regulated pathway for the uptake of cholesterol is disrupted in individuals who inherit defective genes encoding LDL receptor proteins. The resulting high levels of blood cholesterol predispose these individuals to develop atherosclerosis prematurely, and many die at an early age of heart attacks resulting from coronary artery disease. In some cases, the receptor is lacking altogether. In others, the receptors are defective—in either the extracellular binding site for LDL or the intracellular binding site that attaches the receptor to the coat of a clathrin-coated pit (see ). In the latter case, normal numbers of LDL-binding receptor proteins are present, but they fail to become localized in the clathrin-coated regions of the plasma membrane. Although LDL binds to the surface of these mutant cells, it is not internalized, directly demonstrating the importance of clathrin-coated pits in the receptor-mediated endocytosis of cholesterol.
More than 25 different receptors are known to participate in receptor-mediated endocytosis of different types of molecules, and they all apparently use the same clathrin-coated-pit pathway. Many of these receptors, like the LDL receptor, enter coated pits irrespective of whether they have bound their specific ligands. Others enter preferentially when bound to a specific ligand, suggesting that a ligand-induced conformational change is required for them to activate the signal sequence that guides them into the pits. Since most plasma membrane proteins fail to become concentrated in clathrin-coated pits, the pits must function as molecular filters, preferentially collecting certain plasma membrane proteins (receptors) over others.
Signal peptides guide transmembrane proteins into clathrin-coated pits by binding to the adaptins. Despite a common function, their amino acid sequences vary. A common endocytosis signal consists of only four amino acids Y-X-X-Ψ, where Y is tyrosine, X any polar amino acid, and Ψ a hydrophobic amino acid. This short peptide, which is shared by many receptors, binds directly to one of the adaptins in clathrin-coated pits. By contrast, the cytosolic tail of the LDL receptor contains a unique signal (Asn-Pro-Val-Tyr) that apparently binds to the same adaptin protein.
Electron-microscope studies of cultured cells exposed simultaneously to different labeled ligands demonstrate that many kinds of receptors can cluster in the same coated pit. The plasma membrane of one clathrin-coated pit can probably accommodate up to 1000 receptors of assorted varieties. Although all of the receptor-ligand complexes that use this endocytic pathway are apparently delivered to the same endosomal compartment, the subsequent fates of the endocytosed molecules vary, as we discuss next.
Endocytosed Materials That Are Not Retrieved From Endosomes End Up in Lysosomes
The endosomal compartments of a cell can be complex. They can be made visible in the electron microscope by adding a readily detectable tracer molecule, such as the enzyme peroxidase, to the extracellular medium and leaving the cells for various lengths of time to take it up by endocytosis. The distribution of the molecule after its uptake reveals the endosomal compartments as a set of heterogeneous, membrane-enclosed tubes extending from the periphery of the cell to the perinuclear region, where it is often close to the Golgi apparatus. Two sequential sets of endosomes can be readily distinguished in such labeling experiments. The tracer molecule appears within a minute or so in early endosomes, just beneath the plasma membrane. After 5–15 minutes, it moves to late endosomes, close to the Golgi apparatus and near the nucleus. Early and late endosomes differ in their protein compositions; they are associated with different Rab proteins, for example.
As mentioned earlier, the interior of the endosomal compartment is kept acidic (pH ~6) by a vacuolar H+ ATPase in the endosomal membrane that pumps H+ into the lumen from the cytosol. In general, later endosomes are more acidic than early endosomes. This acidic environment has a crucial role in the function of these organelles.
We have already seen how endocytosed materials that reach the late endosomes become mixed with newly synthesized acid hydrolases and end up being degraded in lysosomes. Many molecules, however, are specifically diverted from this journey to destruction. They are recycled instead from the early endosomes back to the plasma membrane via transport vesicles. Only molecules that are not retrieved from endosomes in this way are delivered to lysosomes for degradation.
Specific Proteins Are Removed from Early Endosomes and Returned to the Plasma Membrane
The early endosomes form a compartment that acts as the main sorting station in the endocytic pathway, just as the cis and trans Golgi networks serve this function in the biosynthetic-secretory pathway. In the acidic environment of the early endosome, many internalized receptor proteins change their conformation and release their ligand, just as the M6P receptors unload their cargo of acid hydrolases in the even more acidic late endosomes. Those endocytosed ligands that dissociate from their receptors in the early endosome are usually doomed to destruction in lysosomes, along with the other soluble contents of the endosome. Some other endocytosed ligands, however, remain bound to their receptors, and thereby share the fate of the receptors.
The fates of the receptor proteins—and of any ligands remaining bound to them—vary according to the specific type of receptor. (1) Most receptors are recycled and return to the same plasma membrane domain from which they came; (2) some proceed to a different domain of the plasma membrane, thereby mediating a process called transcytosis; and (3) some progress to lysosomes, where they are degraded ().
Possible fates for transmembrane receptor proteins that have been endocytosed. Three pathways from the endosomal compartment in an epithelial cell are shown. Retrieved receptors are returned (1) to the same plasma membrane domain from which they came (more...)
The LDL receptor follows the first pathway. It dissociates from its ligand LDL in the early endosome and is recycled to the plasma membrane for reuse, leaving the discharged LDL to be carried to lysosomes (). The recycling vesicles bud from long, narrow tubules that extend from the early endosomes. It is likely that the geometry of these tubules helps the sorting process. Because tubules have a large membrane area enclosing a small volume, membrane proteins tend to accumulate there. Transport vesicles that return material to the plasma membrane begin budding from the tubules, but tubular portions of the early endosome also pinch off and fuse with one another to form recycling endosomes, a way-station for the traffic between the early endosomes and the plasma membrane. During this process, the tubules and then the recycling endosome continuously shed vesicles that return to the plasma membrane.
The receptor-mediated endocytosis of LDL. Note that the LDL dissociates from its receptors in the acidic environment of the endosome. After a number of steps (see Figure 13-48), the LDL ends up in lysosomes, where it is degraded to release free cholesterol. (more...)
The transferrin receptor follows a similar recycling pathway, but it also recycles its ligand. Transferrin is a soluble protein that carries iron in the blood. Cell-surface transferrin receptors deliver transferrin with its bound iron to early endosomes by receptor-mediated endocytosis. The low pH in the endosome induces transferrin to release its bound iron, but the iron-free transferrin itself (called apotransferrin) remains bound to its receptor. The receptor-apotransferrin complex enters the tubular extensions of the early endosome and from there is recycled back to the plasma membrane (). When the apotransferrin returns to the neutral pH of the extracellular fluid, it dissociates from the receptor and is thereby freed to pick up more iron and begin the cycle again. Thus, transferrin shuttles back and forth between the extracellular fluid and the endosomal compartment, avoiding lysosomes and delivering the iron that cells need to grow to the cell interior.
Sorting of membrane proteins in the endocytic pathway. Transferrin receptors mediate nutrient uptake and constitutively cycle between endosomes and the plasma membrane. By contrast, opioid receptors are signaling receptors that—after ligand binding—are (more...)
The second pathway that endocytosed receptors can follow from endosomes is taken both by opioid receptors (see ) and by the receptor that binds epidermal growth factor (EGF). EGF is a small, extracellular signal protein that stimulates epidermal and various other cells to divide. Unlike LDL receptors, EGF receptors accumulate in clathrin-coated pits only after binding EGF, and most of them do not recycle but are degraded in lysosomes, along with the ingested EGF. EGF binding therefore first activates intracellular signaling pathways and then leads to a decrease in the concentration of EGF receptors on the cell surface, a process called receptor down-regulation that reduces the cell's subsequent sensitivity to EGF (discussed in Chapter 15).
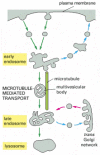
Multivesicular Bodies Form on the Pathway to Late Endosomes
It is still uncertain how endocytosed molecules move from the early to the late endosomal compartment so as to end up in lysosomes. A current view is that portions of the early endosomes migrate slowly along microtubules toward the cell interior, shedding tubules of material to be recycled to the plasma membrane. The migrating endosomes enclose large amounts of invaginated membrane and internally pinched-off vesicles and are therefore called multivesicular bodies (). It is unknown whether multivesicular bodies eventually fuse with a late endosomal compartment or if they fuse instead with each other to become late endosomes. At the end of this pathway, the late endosomes are converted to lysosomes as a result of their fusion with hydrolase-bearing transport vesicles from the trans Golgi network and their increased acidification ().
Electron micrograph of a multivesicular body in a plant cell. The large amount of internal membrane will be delivered to the vacuole, the plant equivalent of the lysosome, for digestion.
The endocytic pathway from the plasma membrane to lysosomes. Maturation from early to late endosomes occurs through the formation of multivesicular bodies, which contain large amounts of invaginated membrane and internal vesicles (hence their name). These (more...)
The multivesicular bodies carry specific endocytosed membrane proteins that are to be degraded but exclude others that are to be recycled. As part of the protein-sorting process, specific proteins—for example, the occupied EGF receptor described previously—selectively partition to the invaginating membrane of the multivesicular bodies (). In this way, the receptors, as well as any signaling proteins strongly bound to them, are rendered fully accessible to the digestive enzymes that will degrade them (see ).
The sequestration of endocytosed proteins into internal membranes of multivesicular bodies. Eventually, all of the internal membranes produced by the invaginations shown are digested by proteases and lipases in lysosomes. The invagination is essential (more...)
Membrane proteins that are sorted into the internal membrane vesicles of a multivesicular body are first covalently modified with the small protein ubiquitin. Unlike multi-ubiquitylation which typically targets substrate proteins for degradation in proteasomes (discussed in Chapter 6), ubiquitin tagging for sorting into the internal membrane vesicles of a multivesicular body requires the addition of only a single ubiquitin molecule that is added to activated receptors while still at the plasma membrane. The ubiquitin tag facilitates the uptake of the receptors into endocytic vesicles and is then recognized again by proteins that mediate the sorting process into the internal membrane vesicles of multivesicular bodies. In addition, membrane invagination in multivesicular bodies is regulated by a lipid kinase that phosphorylates phosphatidylinositol. The phosphorylated head groups of these lipids are thought to serve as docking sites for the proteins that mediate the invagination process. Local modification of lipid molecules is thus another way in which specific membrane patches can be induced to change shape and destiny.
In addition to endocytosed membrane proteins, multivesicular bodies also contain most of the soluble content of early endosomes destined for digestion in lysosomes.
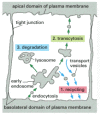
Macromolecules Can Be Transferred Across Epithelial Cell Sheets by Transcytosis
Some receptors on the surface of polarized epithelial cells transfer specific macromolecules from one extracellular space to another by transcytosis (). These receptors are endocytosed and then follow a pathway from endosomes to a different plasma membrane domain (see ). A newborn rat, for example, obtains antibodies from its mother's milk (which help protect it against infection) by transporting them across the epithelium of its gut. The lumen of the gut is acidic, and, at this low pH, the antibodies in the milk bind to specific receptors on the apical (absorptive) surface of the gut epithelial cells. The receptor-antibody complexes are internalized via clathrin-coated pits and vesicles and are delivered to early endosomes. The complexes remain intact and are retrieved in transport vesicles that bud from the early endosome and subsequently fuse with the basolateral domain of the plasma membrane. On exposure to the neutral pH of the extracellular fluid that bathes the basolateral surface of the cells, the antibodies dissociate from their receptors and eventually enter the newborn's bloodstream.
Transcytosis. Recycling endosomes form a way-station on the transcytotic pathway. In the example shown here, an antibody receptor on a gut epithelial cell binds antibody and is endocytosed, eventually carrying the antibody to the basolateral plasma membrane (more...)
The transcytotic pathway from the early endosome to the plasma membrane is not direct. The receptors first move from the early endosome to an intermediate endosomal compartment, the recycling endosome described previously (see ). The variety of pathways that different receptors follow from endosomes implies that, in addition to binding sites for their ligands and binding sites for coated pits, many receptors also possess sorting signals that guide them into the appropriate type of transport vesicle leaving the endosome and thereby to the appropriate target membrane in the cell.
A unique property of a recycling endosomes is that the exit of membrane proteins from the compartment can be regulated. Thus, cells can adjust the flux of proteins through the transcytotic pathway according to need. Although the mechanism of regulation is uncertain, it allows recycling endosomes an important role in adjusting the concentration of specific plasma membrane proteins. Fat cells and muscle cells, for example, contain large intracellular pools of the glucose transporters that are responsible for the uptake of glucose across the plasma membrane. These proteins are stored in specialized recycling endosomes until the cell is stimulated by the hormone insulin to increase its rate of glucose uptake. Then transport vesicles bud from the recycling endosome and deliver large numbers of glucose transporters to the plasma membrane, thereby greatly increasing the rate of glucose uptake into the cell ().
Storage of plasma membrane proteins in recycling endosomes. Recycling endosomes can serve as an intracellular pool for specialized plasma membrane proteins, enabling them to be mobilized when needed. In the example shown here, insulin binding to the insulin (more...)
Epithelial Cells Have Two Distinct Early Endosomal Compartments But a Common Late Endosomal Compartment
In polarized epithelial cells, endocytosis occurs from both the basolateral domain and the apical domain of the plasma membrane. Material endocytosed from either domain first enters an early endosomal compartment that is unique to that domain. This arrangement allows endocytosed receptors to be recycled back to their original membrane domain, unless they contain signals that mark them for transcytosis to the other domain. Molecules endocytosed from either plasma membrane domain that are not retrieved from the early endosomes end up in a common late endosomal compartment near the cell center and are eventually degraded in lysosomes ().
The two distinct early endosomal compartments in an epithelial cell. The basolateral and the apical domains of the plasma membrane communicate with separate early endosomal compartments. But endocytosed molecules from both domains that do not contain (more...)
Whether cells contain a few connected or many unconnected endosomal compartments seems to depend on the cell type and the physiological state of the cell. Like many other membrane-enclosed organelles, endosomes of the same type can readily fuse with one another (an example of homotypic fusion, discussed earlier) to create large continuous endosomes.
Summary
Cells ingest fluid, molecules, and particles by endocytosis, in which localized regions of the plasma membrane invaginate and pinch off to form endocytic vesicles. Many of the endocytosed molecules and particles end up in lysosomes, where they are degraded. Endocytosis occurs both constitutively and as a triggered response to extracellular signals. Endocytosis is so extensive in many cells that a large fraction of the plasma membrane is internalized every hour. To make this possible, most of the plasma membrane components (proteins and lipid) that are endocytosed are continually returned to the cell surface by exocytosis. This large-scale endocytic-exocytic cycle is mediated largely by clathrin-coated pits and vesicles.
Many cell-surface receptors that bind specific extracellular macromolecules become localized in clathrin-coated pits. As a result, they and their ligands are efficiently internalized in clathrin-coated vesicles, a process called receptor-mediated endocytosis. The coated endocytic vesicles rapidly shed their clathrin coats and fuse with early endosomes.
Most of the ligands dissociate from their receptors in the acidic environment of the endosome and eventually end up in lysosomes, while most of the receptors are recycled via transport vesicles back to the cell surface for reuse. But receptor-ligand complexes can follow other pathways from the endosomal compartment. In some cases, both the receptor and the ligand end up being degraded in lysosomes, resulting in receptor down-regulation. In other cases, both are transferred to a different plasma membrane domain, and the ligand is thereby released by exocytosis at a surface of the cell different from that where it originated, a process called transcytosis. The transcytosis pathway includes recycling endosomes, where endocytosed plasma membrane proteins can be stored until they are needed.