

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Institute of Medicine (US) Committee for the Study of Health Consequences of the Stress of Bereavement; Osterweis M, Solomon F, Green M, editors. Bereavement: Reactions, Consequences, and Care. Washington (DC): National Academies Press (US); 1984.

Bereavement: Reactions, Consequences, and Care.
Show details
This chapter reviews the available data on the biologic events that occur during the grieving process and proposes several kinds of pathways through which grief might increase vulnerability to physical illness. The degree to which this vulnerability occurs and results in illness is still open to question.
Research to date has shown that, like many other stressors, grief frequently leads to changes in the endocrine, immune, autonomic nervous, and cardiovascular systems; all of these are fundamentally influenced by brain function and neurotransmitters. However, the significance of these changes is not well understood. They may be primarily adaptive physiologic responses that in some persons become maladaptive and physiologically deleterious.
The notion of a normal adaptive response becoming harmful is not new, especially in the context of the immune system. For instance, there is evidence that chronic active hepatitis is a virally induced or drug-induced autoimmune disease. Craddock 16 suggested other ways the body's natural defenses may cause disease: the appropriate response to endotoxemia is fever and leukocytosis, but if the response is increased it may result in shock, consumption coagulopathy, tissue necrosis, hemorrhage, and death. The complexing of soluble antigen with antibody is a part of the immune response, but deposition of too many immune complexes can lead to severe vascular disease. Likewise, lymphatic hyperplasia is necessary to combat the Epstein-Barr virus, but in severe infectious mononucleosis it can cause serious tissue destruction.
The hypothesis that a normal adaptation to grief can become unregulated and lead to illness is considered further in the section of this chapter on a "psychoneuroimmunoendocrine system." This view is consistent with contemporary stress theory, 17 in which it is postulated that a stressor (x) produces certain transient biological or psychosocial reactions (y) that may (or may not) cumulatively lead to certain health consequences (z). Reactions and consequences may be modified by a number of factors, and the consequences may themselves activate other stressors. In simplified form, the stress model looks like this:
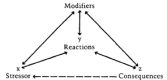
FIGURE 1
SOURCE: Adapted from Elliot and Eisdorfer.
Epidemiologic studies linking the stress of bereavement with adverse consequences to physical and mental health have been reviewed in Chapter 2; these are x-z correlations, using this model. This chapter reviews the many studies of prompt biological reactions to the stress of bereavement (x-y) and presents conceptualizations and promising leads—but not confirming data—on the implications of these particular physiologic changes for enduring health consequences (x-y-z) in human beings.
ANIMAL MODELS
Finding out more about the involvement of complex biological systems in grief would be greatly facilitated if animal models were available. The history of medical investigation is testimony to the importance of animal research in understanding the pathophysiology of human disease.
Animal models rarely provide precise replicas of human conditions because of species differences. But nature, in evolution, has been conservative, and human beings share many physiological and psychological processes with other species. Thus, a number of the studies discussed in this chapter are based on research with monkeys, dogs, or rats. Social attachments, for example, are widely represented in mammals and in birds, particularly between the young and their social companions, parents, and siblings. Acute behavioral responses of infants to separation from their mothers are quite similar across species, including rat, guinea pig, cat, dog, monkey, and human being. And the few chronic biological responses to early separation that have been studied show striking similarities between monkeys and rats: a similar reduction in rapid eye movement (REM) sleep with insomnia, a lowering of cardiac rate, a thermoregulatory disturbance with decrease of core temperature, and a (very recently) demonstrated decrease in immune competence. 1,26,27,30,46,62,63
What do these findings indicate about the biology of human grief? Because of the differences between immature and mature organisms, between separation and permanent loss, and between various mammalian species, they can only be suggestive. They confirm that the response to separation has deep evolutionary roots and some biological impact on systems controlling vital processes of sleeping, circulation, thermoregulation, and immune surveillance. These studies require constant innovation in developing methods to detect physiologic changes, methods that may one day be applicable to human beings.
Most important, animal studies provide new conceptual models for approaching an understanding of human bereavement. Examples are given in the section on social relationships as biological regulators and in Chapter 7. In an additional example, a single aspect of grief can be isolated and studied in terms of its biological substrate, as in the work of Weiss 83,84 on learned helplessness. The helplessness of human grief is surely different from the state induced in rats by inescapable shock. Yet an understanding of the simpler case should provide clues about what to measure and how to begin to study the much more complex interrelationship of affects and biological changes that occur in human grief.
BIOLOGIC STUDIES OF BEREAVED HUMAN BEINGS
There have been very few studies employing biologic variables in actively grieving people. The first study of this sort was Lindemann's, 48 in which he concentrated on what he termed the "syndrome" of acute grief. The data he collected from the relatives of victims of the "Coconut Grove" nightclub fire in Boston in the early 1940s, as amplified and extended in the work of Parkes 59 in London in the late 1960s and more recently in studies by Clayton 13 and others (e.g., Bartrop et al. 5), give a picture of the behavioral and physiologic symptomatology of grief, as outlined in Table 1.
TABLE 1
Adult Grief Responses (Human Studies).
The symptoms described in Table 1 are present to a greater or lesser extent in most bereaved people for a period of weeks or months and in brief reactivation bouts, such as anniversaries, for years. They are the best evidence, thus far, that bereavement involves a physiologic disturbance. Unfortunately, symptoms such as these are not infallible indicators of physiologic change, nor are most of them very specific indicators of which organ or system may be involved. More specific physiologic data have been collected in animal studies of mother-infant separation; and these are summarized in Table 2 (page 165).
TABLE 2
Infant Separation Responses (Animal Studies).
In 1944, Lindemann described having made respiratory tracings by polygraph of bereaved people and discovered an unusual pattern that he indicated would be described in detail in a subsequent paper. That paper was never written, and the idea dropped from sight until 1980 when Schiffman and his colleagues, 70 looking for a heritable respiratory defect in relatives of Sudden Infant Death Syndrome (SIDS) victims, found compromised ventilation in recently bereaved parents of children lost to SIDS, suggesting that the chemical regulation of their breathing might be defective. It is unclear from this study whether the alteration in the autonomic regulation of breathing is genetic or acquired, and if acquired whether it is related to the grief state. In order to understand this more fully, Shannon and his colleagues at the Massachusetts General Hospital in Boston are conducting a study to determine the relationship between grieving, depression, and autonomic regulation of breathing and cardiovascular function in recently bereaved parents of SIDS victims, parents whose child died from other causes, depressed patients, and nongrieving/nondepressed parents (D. Shannon, personal communication).
The pituitary-adrenocortical system was studied in the parents of children with leukemia, both before the child's death 86 and in the period of bereavement. 33,34 The researchers found that the operation of psychological defenses played a major role in modifying the extent of activation of this system during the threat of loss. A subject's level of cortisol (a hormone secreted by the adrenal cortex in response to adrenocorticotrophic hormone [ACTH] from the pituitary) could be predicted on the basis of an assessment of defense effectiveness. In follow-up visits six months and two years after the child's death, however, parents no longer maintained the same rank order of cortisol levels as they had prior to the death; cortisol levels varied directly with the extent of active grieving (at the six-month follow-up visit) as judged by an assessment procedure carried out without knowledge of the hormonal data. From these data, it might appear that grief is associated with adrenocortical activation, which continues as long as grief is an ongoing conscious psychological process. However, in those who had been actively grieving at six months and whose grief appeared to have run its course by the two-year return visit, cortisol levels remained high and unchanged. This pioneering study raised as many questions as it answered and gave an early indication of the complications that may arise in this kind of research.
Current psychoendocrine studies at Yale measure plasma levels of pituitary prolactin and growth hormone, as well as cortisol, in recently bereaved subjects during interviews in which the loss experiences were reviewed. Those whose growth hormone output increased during the interview tended to be subjects whose distress had been worsening in the two months between the loss and the interview. Measures of distress during the interview did not correlate in a simple way with pitui tary hormone responses, but when one takes into account subtle aspects of the interview situation and the subject's coping mechanisms, interesting correlations begin to emerge. 35,39,41,52 The data suggest a specificity between certain psychological variables and certain hormones; a selective organization of neurohumoral responsiveness seems to be involved in the independent regulation of prolactin, growth hormone, and ACTH.
In the past few years, two studies have addressed the possibility that bereaved individuals have abnormal responses of their cell-mediated immune system. This research was stimulated by suggestions that malignancies may originate more readily among the bereaved. Although, as discussed in Chapter 2, the association is not firmly established on epidemiologic grounds, the clinical observations are frequently intriguing. Researchers in Australia 5 and the United States 7l have found that bereaved persons have an impaired function of T-lymphocytes, the agents of cell-mediated immunity. The lessened T-cell function was not a concomitant of changes in number of circulating T-cells, or of changes in adrenal or thyroid hormones in blood plasma. Neither of these studies was designed to tell whether the reduced T-cell function posed a clinically significant vulnerability of the subjects to disease. These studies require follow-ups that will include investigations of the mounting of specific immune responses. Such research would permit consideration of the relationship of immune function to bereavement and the possible relevance to clinical vulnerability.
PROMISING RESEARCH APPROACHES
Much less is known about the physiology of human bereavement than, for example, about the physiology of exercise or pregnancy. Thus, the most important statement that can be made about the biology of grieving relates to the need for research in this area.
There are at least four areas of knowledge that give useful clues about where to begin: the symptomatology of grief, the epidemiology of bereavement, recent findings in the biology of depression and anxiety disorders, and current knowledge about neuroregulatory mechanisms. In these areas the autonomic, physiologic, biochemical, and endocrinologic systems most likely to be affected by grief and the bereavement process can be identified—regardless of whether the response is viewed as a subcategory of stress response, as a withdrawal of psychobiological regulators, as an adaptive response to a natural event, or as the activation of a neurobiological substrate.
Changes in these systems over time should be studied to reveal the processes of recovery from bereavement as well as the form and duration of the acute disruption. How do children and aging people differ in grief from young and middle-aged adults? What are the special features of the biological responses that are relevant for adverse consequences? In all these areas, the psychological and behavioral responses will have to be related to the biological changes, because there is powerful interplay between these systems in determining the final outcome.
Clues from the Symptomatology of Bereavement
Research should take into account the natural division of symptoms into acute waves of distress, lasting minutes at a time, and chronic disturbance, which can last weeks and months. Acute waves of distress will be reasonably easy to study, because in many people they can be precipitated by an empathetic interviewer and their elicitation is often viewed as helpful to the subject. The symptoms of sighing respiration, dyspnea, substernal tightness, palpitation, weakness, and crying suggest that tests should be made of respiratory control and blood gases, autonomic function (particularly in the cardiovascular system), and energy metabolism (including the rapidly responding hormone systems of the pituitary, thyroid, and adrenal glands). These measures will be most useful if evidence about the subject's inner affective and cognitive state is collected simultaneously and integrated with the physiologic data, so that a comprehensive view of the psychobiological organization of the acute distress "waves" will be obtained.
This basic knowledge will be useful in planning clinical studies of patients at risk because of preexisting disease such as asthma, pulmonary emphysema, coronary heart disease, or congestive heart failure. These conditions are most likely to be affected by the physiologic changes of acute distress waves.
The chronic form of disturbance is characterized by symptoms involving a number of different biological systems that are regulated or powerfully influenced by the central nervous system through its autonomic, neuroendocrine, and musculoskeletal outflow channels. Many of the symptoms may be related to the neural regulation of the sleep-wake cycle and related biological rhythms, that is, to disturbance of chronobiological organization. The insomnia, chronic fatigue, restlessness, appetite disturbance, cognitive and perceptual disturbance, and even the illusions and depressive affect may be manifestations of altered integration or patterning of biological rhythms. If this is true, circadian rhythms of body temperature, physical activity, cardiac rate, urinary output, and other vital functions should be examined for evidence of free running (non-24-hour rhythmicity) or internal desynchronization (lack of consistent relationship between physiologic systems).
Clues from the Epidemiology of Bereavement
As documented in Chapter 2, a number of studies suggest that bereavement may enhance vulnerability to neoplastic disease, infection, cardiovascular disease, substance abuse, or depression.
The existence of hormone-sensitive cancers raises the possibility that alterations in hormonal milieu following bereavement may have clinical importance. Estrogen-dependent breast cancers and testosterone dependent prostatic cancers are some of the best known examples of this sort. Studies are needed of ovarian, testicular, and adrenal hormonal regulation in bereavement, and of the hypothalamic-pituitary stimulating hormones that control these target glands. The potential involvement of the immune system in the recognition and suppression of neoplasia makes it an obvious subject of study during grief. Important interactions are already known among adrenal cortical hormones, autonomic nervous system neurotransmitters, and the immune system.
Susceptibility to bacterial and viral infections has been linked to bereavement in some studies ( Chapter 2). This, too, suggests a need for a combined endocrine and immune system investigation. Resistance to infectious agents is known to depend critically on hormonal milieu and on humoral and cell-mediated immune mechanisms. The incidence of pneumonia as a terminal event in the bereaved might also indicate disturbed respiratory regulation during grieving.
The link between disease and bereavement is strongest for the cardiovascular system. Sudden cardiac death, cardiac arrhythmias, myocardial infarction, and congestive heart failure are the most frequently mentioned conditions of that system associated with grief. 72 This presents us with the clue that disturbances in autonomic cardiovascular regulation and in circulating catecholamines may be present in bereavement and may be exaggerated in patients with preexisting cardiovascular disease of the arteriosclerotic or hypertensive variety. Studies have shown that patients with congestive heart failure 12 and with essential hypertension 82 and arrhythmias 60,61 are particularly prone to exacerbation of their condition in response to threatened or actual loss of human relationships. Enough is known about the pathophysiology of these conditions to make quite specific hypotheses as to the probable autonomic and neurohumoral mechanisms involved. In addition, methods to test such hypotheses are currently available.
Vulnerability of the bereaved to substance abuse suggests that they are self-medicating themselves in an effort to alleviate their psychic pain and bodily discomfort. The possibility that sensory and pain thresholds are altered in the bereaved has not been systematically evaluated by modern methods, nor have any of the newer approaches to the study of endogenous, opiate-like peptides and their receptors been employed with this group. Alternatively, substance abuse may be related to depression, a possibility discussed in the next section.
Clues from the Endocrinology and Biochemistry of Depression and Anxiety States
Two well-delineated mental disorders, major depression and panic disorder, may share a common neurobiological substrate with the response to loss in the bereaved. This hypothesis bears testing by looking for the characteristic biochemical, physiologic, and pharmacologic responses of these two patient groups in adults following bereavement uncomplicated by diagnosed psychiatric illness.
Endocrine control during depressive illness and recovery still requires fuller study and documentation. Abnormalities of hypothalamic neurotransmitter systems controlling the pituitary output of ACTH, luteinizing hormone, and prolactin have been found in depressed patients; these findings may also reflect abnormalities in neurotransmitters in other areas of the brain that mediate mood and behavior. Are similar abnormalities found in the bereaved? How are these related to peptide levels in plasma, to immune function, to psychological status?
A characteristic change in sleep-wake state regulation (a shortened latency to the first REM period) has been found in many depressed patients, and reverts to normal with easing of the illness. Might a similar abnormality be found in disturbed sleep following bereavement?
Patients with panic disorder show two biological abnormalities that may have some relevance to the psychobiology of acute grief. They have abnormally wide fluctuations in certain autonomic variables (particularly skin resistance), especially in the prodromal period before a panic attack. 45 And they are extremely susceptible to precipitation of a panic attack by intravenous lactate infusion. 37 This susceptibility appears to be blocked by the tricyclic antidepressant drug, imipramine. The waves of distress, a hallmark of acute grief, bear some resemblance to acute panic attacks; can they be precipitated by lactate infusion and are they heralded by similar patterns of autonomic instability?
The complexity of interactions between the psychologic and the physiologic has been illustrated in a study by Kraemer and McKinney. 42 They used drugs to alter the function of two neurotransmitters that have been implicated in the genesis of depression. With drug doses that failed to produce any detectable effects in juvenile monkeys while they were in their home-peer groups (thus obviating any spurious side effects), increases or decreases in behavior patterns of despair could be produced following peer separation. Thus, only when beset by the significant psychological stress imposed by loss of their peers were these animals responsive to the drug regimen. This complex interaction, in which events at the psychological level affect physiologic processes and where the physiology in turn alters psychological functioning, is a ubiquitous quality of primate psychobiology.
Two neurochemical changes have been found in rhesus monkey infants separated from their mothers and placed in isolation: increases in catecholamine-synthesizing enzymes and in hypothalamic levels of the neurotransmitter serotonin. 8 Suomi and his colleagues 77 subsequently studied rhesus infants raised in peer groups without their mothers until 90 days and then subjected to repetitive peer separations; the tricyclic antidepressant, imipramine, decreased excessive self-clasping and prevented reduction in play behavior characteristically found after such experiences. However, social contact also was reduced and locomotor behavior increased by the drug. Although neurochemical changes clearly occur in infant monkeys separated from social companions, the neurochemical basis for the behavioral changes and the mechanism of their modification by psychoactive drugs are not yet clear.
Clues from Current Knowledge of Neuroregulatory Mechanisms
The neuroregulatory pathways that are known to mediate the conversion of life experiences into changes of bodily function provide many insights into the biology of grief. Whether some or all of these actually are involved in human grief is an empirical question that has received little study. Research on stress physiology has identified many neural and endocrine control systems that are at least a good place to begin investigation. These involve the autonomic neural system, endocrine events, and immune changes related to endocrine and direct neural influences. 17
Autonomic Neural Effects. Since the time of W. B. Cannon, 9 it has been known that emotional states of rage and fear affect such biological functions as blood flow and cardiac rate by increasing the activity in widely distributed nerve fibers called the autonomic nervous system. This system comprises two main divisions, distinguished by anatomic features, range of distribution, and characteristic neurotransmitter types into sympathetic and parasympathetic divisions. These systems are both afferent and efferent and serve to control and coordinate vital functions of digestion, metabolism, circulation, and respiration. In addition to maintaining what Cannon 10 termed ''homeostasis," the autonomic system mobilizes the resources of the body in preparation for "fight or flight."
Autonomic nervous system function has been found to be altered in a variety of ways during an organism's responses to exercise, sex, mental arithmetic, and relaxation and meditation states. Although there have not as yet been any specific and systematic studies of autonomic neural physiology during grief, a number of studies have found autonomic changes to be associated with themes and events associated with rejection, abandonment, and loss. No clear-cut pattern has emerged, however, and the situations and affects were quite disparate.
Patterns of physiologic response following loss have been studied in the biotelemetry research of Reite and his colleagues. 62,63 Using implanted devices to record a number of measures simultaneously in free moving monkeys, they studied the response of pigtail macaques to infant-mother separation. These data show that after initial increases in heart rate and body temperature in the first day after the loss, sharp drops in both measures occurred during the first night, and levels did not return to normal for the next several days. Disturbances in sleep, particularly in REM sleep, emerged in the first evening and disruption of normal sleep patterns continued throughout the 10-day study. Thus, the period of heightened heart rate and temperature roughly coincided with the protest behavior that was observed, while the sleep disturbance and decrease in the levels of these parameters approximated the subsequent despair phase of the response.
One example will illustrate how knowledge of autonomic nervous system physiology could help researchers understand the biology of bereavement. The excess mortality found in several epidemiologic studies during the first six months after bereavement is predominantly caused by cardiovascular events. And studies by Engel 18 and Reich 60,61 have described a number of episodes of sudden death proved or presumed to be due to ventricular fibrillation that occurred in the setting of loss and grief. How could grief stop a heart?
Working with dogs, Lown and his associates 49 have identified a phase during the electrical events of cardiac contraction, lasting only micro seconds, when the heart is particularly vulnerable to developing multiple beats and even ventricular fibrillation in response to a weak electrical current. Verrier, 80 working with Lown, then studied the threshold for this "ventricular vulnerable period" as it might be influenced by the emotional state of the experimental animal and by brief episodes of coronary narrowing and reperfusion (as might occur with coronary artery spasm). They discovered that unfamiliarity with the testing chamber and expectation of receiving mild punishment reduced the threshold by 41 percent. They were then able to analyze this effect in terms of the contribution of sympathetic and parasympathetic autonomic influences. Using pharmacologic blocking agents, measurement of plasma norepinephrine and epinephrine, and surgical excision of the stellate ganglia, they found that the reduced threshold was the result of increased sympathetic influences—both along neural pathways and through increased circulating levels of norepinephrine and epinephrine.
These experimental findings do not bear directly on the question of how grief may contribute to cardiac arrhythmias, for they involved a different emotional setting and a different species. But they do suggest how growing knowledge about autonomic mechanisms may help direct the search for biological mechanisms in grief that may predispose individuals to health risks.
Endocrine Events. Psychological events cause many changes in the internal hormonal milieu, principally via the hypothalamic-pituitary axis. The response of the hypothalamic-pituitary-adrenocortical axis to stress has been well documented. Increased urinary and plasma levels of 17-hydroxycorticosteroids (17-OHCS) have been demonstrated in response to movies of emotionally charged material, 21 the stress of army basic training, 66 test-taking, aircraft flight, preoperative period novel situations, 22 hospital admission, 53 athletic events, 78 and other stressful situations. 51
Secretion of other hormones also is altered by stressful situations. Testosterone decreases during stress. 43 Sowers and colleagues 74 found that physical and psychological stress related to diagnostic procedures and surgery was associated with decreased thyrotropin and thyroxine and with increased prolactin, growth hormone, cortisol, and luteinizing hormone, with no evident effect on follicle stimulating hormone. Stress increases prolactin in the morning when the level is low, and suppresses it in the afternoon when levels are high. 23 Mason 51 found that in monkeys performing a learned avoidance task, there was an increase in catabolic hormones and growth hormone and a decrease in anabolic hormones.
Different emotional responses to stress are reflected in hormonal patterns. Von Euler et al. 21 measured urinary catecholamines in young men shown films of murders, fights, tortures, executions, and cruelty to animals and found that those who were the most emotionally distressed by the films had the most significant endocrine reaction. Psychiatric interviews, field observations, and measurements of 17-OHCS excretion in army recruits showed that low excretors had better defense mechanisms and that high excretors were less successful in dealing with the stress of basic training. 66
Animal studies have shown that stress, especially uncontrollable or inescapable shock, causes many changes in norepinephrine, epinephrine, and dopamine concentrations in peripheral blood and in different parts of the brainstem, hypothalamus, and limbic system. 68 Although the link between these neuroendocrine substances and the hypothalamic-pituitary axis has not been completely elucidated, a relationship certainly exists. The "helpless-hopeless syndrome" seen in animal studies of inescapable shock situations appears to have special effects on endocrine activity that may have some potential relevance for studies of human stress, including bereavement. In patients entering the psychiatric ward of a general hospital, most of whom had recently suffered a very difficult interpersonal event, Board et al. 7 found that those diagnosed as the most distressed had the highest levels of 17-OHCS and that they tended to be sad, hopeless, and "retarded" rather than agitated. In animal studies, investigators have found that stress-induced increases in plasma and urinary 17-OHCS are highest in situations in which long-established rules have changed and previously effective behavior no longer works to alleviate the stress. 21
Research on the response of the endocrine system to disruption of attachments has been conducted by Levine and his associates in both the squirrel monkey and the rhesus macaque. 15,47 These studies generally have involved relatively brief and often repeated separation experiences, usually lasting for periods of hours or a few days. This work has shown quite consistently in both species that the sudden loss of the partner causes rapid and often dramatic increases in adrenal function with marked cortisol secretion evident even 30 minutes after separation. In general, the period of highest cortisol concentration is found during the first 24 hours following separation, with a decline thereafter. This period coincides with the initial phase of "protest" in response to loss. In squirrel monkeys, at least, both the mother and infant show these rapid cortisol increases; the mother's return to baseline levels is more rapid than her infant's. 14 The diminution of response after the first day not withstanding, recent research by this group reveals that increased adrenal activity may be detected as long as two weeks after the separation. 85
Further research 25,81 illustrates another significant fact that has emerged in physiologic research—the potential for discontinuity between the behavioral and physiologic measures of the loss response. Consider the following: infant monkeys can develop strong attachment bonds toward inanimate surrogate mothers. When infant squirrel monkeys are separated from their surrogate mothers, they show a very dramatic behavioral response of the "protest" variety. If the infant, at separation, is removed to a novel pen, it shows a large increase in blood cortisol—thus, the behavioral measures of stress used here, that is, screaming and agitated activity, are congruent with the physiologic measure. If the infant is left in its home cage and it is the surrogate that is removed, however, the infant still screeches, but its cortisol level remains undisturbed. In rhesus monkeys separated from their biological mothers, elevated cortisol coincided with the protest pattern of the first day, but cortisol levels returned to baseline after 24 hours while various aspects of the initial behavioral response continued for 11 days after separation. 15
Immune Changes Related to Endocrine Events. It is well known that adrenal cortical activity has the effect of reducing circulating lymphocytes and increasing thymic involution, resulting in diminished immune function. 55,58 Riley and others have conjectured that an increase in adrenal cortical activity induced by stressful manipulation of experimental animals was causally related to an enhanced susceptibility to the growth of malignant tumors. 64,65 Thymic involution and fewer circulating lymphocytes implicate T-cell deficiency as a possible factor in tumor susceptibility; hence Riley and others have shown tumor enhancement. But Newberry 57 and others have found the opposite effect: that some stressors suppress tumor development, which suggests that timing, tumor model, and type of stressor all are factors to be considered.
Immune Changes Related to Psychological Events: Psychoneuroimmune Reactions. Palmblad 58 demonstrated that stress-induced immunosuppression occurs independent of endocrine function. Based on previous findings that the surgical stress of thoracotomy increased the number of lung metastases and tumor growth, Hattori and colleagues 24 administered immunopotentiators (streptococcal preparation and mitogens) to animals with cancer and reduced the number of metastases after thoracotomy, suggesting that tumor spread after surgery was due to immunosuppression.
Coe et al. 15 report that squirrel monkeys separated from their mothers for seven days showed immunosuppression as well as "a marked reduction in their antibody response" when challenged with a benign bacteriophage. However, in keeping with the importance of the environment in affecting response to loss, infants left in their home cage during separation showed less immunologic deficit than those moved to novel settings. In support of hypotheses regarding the role of genetic factors, parallel studies in the rhesus macaque have suggested rather different results. Despite their typically pronounced behavioral response to separation, but perhaps related to their lower adrenal response profiles, the rhesus failed to show immunosuppression after seven days of separation.
The development of virally induced neoplasms in animals and infectious diseases in human beings and animals may be enhanced or diminished depending on the type of stressful situation. 2-4,17 Stress has been shown to affect humoral and cell-mediated immunity and macrophages, with acute stress increasing the immune response and chronic stress decreasing it. 11,75 Stress such as sleep deprivation has caused initial immunosuppression, followed by enhancement of the immune response. 58
In reviewing the positive and negative effects of stress on immune function in animals, Ader 3 concluded that "in general, high stress scale scores combined with a presumed unsuccessful coping response were correlated with depressed immunologic defenses." The impact of stress on disease depends on "the quality and quantity of stressful stimulation, the quality and quantity of immunogenic stimulation, the myriad host factors upon which stress and immunogenesis are superimposed, the temporal relationship between stress and immunologic stimulation, procedural factors such as the nature of the dependent variable and sampling procedures, and the interaction between any or all of the above."
Human beings also respond to stress with immunologic changes, which probably are determined by a combination of the same variables. Among psychiatry trainees facing their final exam for fellowships, those who were highly stressed psychologically displayed transient increases in T- and B-cells but decreases in plaque formation and response to mitogens prior to the exam. These values returned to normal later. Palmblad 58 cites studies of bereavement and other life changes that have shown depression of both humoral and cell-mediated immunity, correlating most highly with the amount of subjectively perceived stress. Schleifer and his colleagues 71 studied immune function in a group of 15 widowers and found that in vitro lymphocyte response to stimulation by common mitogens was significantly lower in the two months following the deaths of their wives, compared with prebereavement levels. The total numbers of B- and T-cells were unaltered, suggesting alterations in subpopulation ratios or defects in responsivity. After two months, some responses improved but others did not. Bartrop and others5 also found that T-cell function was significantly decreased after bereavement, without a change in T-cell number. There was no change in B-cell function, nor in adrenocortical, adrenomedullary, or thyroid hormones.
Further evidence of psychoneurologic influence over the immune system is provided by the many studies of behavioral conditioning of immune function. 4 Ader 2 has written a historical account of conditioned immunobiologic responses, in which he notes that studies of such responses originally done in Russia early in this century were based on the belief that immunologic mechanisms were basically physiologic phenomena under complete control of the brain. Repeated experiments have shown that so-called "natural" defense mechanisms (cell-mediated immunity, phagocytosis, etc.) were readily conditionable, with less evidence being provided for conditionability of humoral immunity. Ader points out the variability of results, depending on the antigen used, the conditioned stimulus, and other factors, and states that it is too early to determine the mechanism by which conditioned immunosuppression occurs. There is also evidence that immunosuppression can be hypnotically induced in human beings. 6,73,75
These findings indicate that immune systems are influenced directly by the central nervous system as well as by hormonal factors. The precise mechanisms are as yet unclear, but it appears that stress of many kinds affects both the endocrine and immune systems and that grieving, considered as a type of stress, would be expected to do so as well.
A cautionary note about interpreting measurements of immune function must be added, however, along with an argument for clinically relevant research involving human subjects. Assessment of T-cell or B-cell functions in vitro is of uncertain meaning in regard to actual vulnerability to infection. When these measures are done in a clinical setting, extreme values generally do indicate immune incompetence. Further evidence is required to demonstrate that the association of grieving with lesser immune changes actually bears a relationship to health.
Studies of bereavement and immune function have primarily investigated mitogen-induced lymphocyte stimulation, which is an in vitro correlate of immunity and provides a general measure of the lymphocyte's ability to synthesize deoxyribonucleic acid and ultimately divide.
Mitogen activation is a useful probe, but it does not involve processes associated with the induction of an immune response. The ability to mount an immune response to an antigen is a primary and specific function of the immune system requiring both antigen recognition and effector processes. One approach, which has yet to be applied systematically, is the direct measure of immune functions in human beings in vivo. For example, by skin testing two agents to which nearly everyone is sensitive ("allergic"), such as candida or trichophyton, it seems possible to establish the nature and intensity of the skin response as a function of psychological state, changing over time. Alterations in measurements of antibodies to cytomegalovirus or herpes virus might also be considered. Another potentially helpful advance would be development of reliable techniques for assessing the incidence of minor cutaneous infections, head colds, and other infections during the grieving process. There are not only urgent needs but also promising opportunities for the direct study of the clinically relevant biology of grieving, including immunity, in the human being.
GRIEF AS AN ADAPTATION IN A "PSYCHONEUROIMMUNOENDOCRINE SYSTEM"
There are abundant data on the effect of psychological events on the endocrine and immune systems and a growing body of information on the endocrine-immune interaction. Probably these interactions are not a one-way street. Thus, a "psychoneuroimmunoendocrine system" can be envisioned. Grief may be viewed as a series of events or reactions in an activated "psychoneuroimmunoendocrine" system that may come to a favorable—or at least neutral—outcome for health or to less favorable and maladaptive consequences.
As shown in Figure 2, the system in simplified form is composed of elements that fluctuate in terms of blood level or activity level. Implicit in the diagram is the existence of effector arms linking each system to another, and also feedback arms. The description in previous sections has already dealt extensively with many of the effectors. There are possibilities for connections with other mechanisms and systems.

FIGURE 2
The Psychoneuroimmunoendocrine System
This system is hypothesized to be a basic, coordinated mechanism for the response of the organism to stress or arousal and to more chronic events such as loss and bereavement. The activated hypothalamus secretes corticotropin releasing factor, which triggers release of a large peptide synthesized in the pituitary. This peptide, pro-opio-melanocortin (POMC) contains ACTH, ß-endorphin, and other peptides with less well understood functions. Some of these peptides or some smaller peptides derived from them are believed to play a role in affective and cognitive function.
There is the possibility that the secretion of one or more products of POMC leads to the activation of various portions of a psychoneuroimmunoendocrine system with separate and important functions. Thus, the effect of adrenal stimulation may be of consequence in the metabolic "preparation" for threat of injury. The catabolic state induced by adrenal activation may provide the organism with an important substrate for energy metabolism. The simultaneous secretion of substances that suppress immune function may serve as a protection by transiently halting the immune response to a flood of antigens appearing from endogenous (proteolysis) or exogenous (injury) effects. The presence of ¬endorphin and other brain-related peptides may provide the transient analgesic or even mental alertness required to focus on the external threat and deal with it in the most appropriate way possible.
This integrated system may also be utilized in dealing with chronic arousal such as grief. In this circumstance, the presence of neurohormones that may assist in denial and may therefore permit the metering of recognition of external events in a slow and staged way may be called upon with each new flood of arousing stimuli. The immune or other effects of the system may become increasingly "maladaptive" in the chronic situation. The adverse biological consequences of continued, "inappropriate" need for activation of the system are evident. Perhaps certain genetic characteristics, the use of culturally learned practices, the intervention of key figures in the social network, or other such factors can reduce a person's "need" to utilize this system in the solution of the problem of grief.
The approach outlined above has the merit of suggesting how immunosuppression, for example, might come about as a maladaptive use of a system, functioning on behalf of the need for unusually prolonged or excessive autopalliation in the face of grief that is particularly difficult to resolve. Such a unified approach, which ties together psychological, endocrine, and immune events, is most attractive but will require validation by much clinical, cross-disciplinary, long-term observation.
CONCEPTUAL FRAMEWORKS
In the absence of good data, theories abound. And theoretical positions determine what questions are asked and what data are collected as well as how studies are interpreted. Thus, it is important to have some insight into the main theoretical outlooks with which clinicians and investigators approach the subject of the biological nature of grief. The remainder of this chapter summarizes three current theoretical frameworks. It should be noted that they are not mutually exclusive and may even be integrated eventually as data are obtained.
Bereavement as a Stressor
As mentioned previously, the current information available on stress and health has been summarized in an Institute of Medicine report. 17 The physiologic responses to a range of external events viewed as stressful have identifiable effects on biological systems. The stress imposed on the bereaved person derives from the internal disequilibrium created both by loss and the attempt to regain homeostasis and by the external disruption of the environment following loss of someone close. Grief could be considered a reaction to stress that is unusual in at least two respects—its chronicity (the loss of the loved object is forever) and the absence of an effective repertoire of behaviors to undo the stressor (the love object cannot be brought back without hallucinations or a denial of reality).
Rose and his colleagues 66 pointed out that successful mechanisms for managing stress are not always equivalent to adaptive coping mechanisms. In bereavement, the common immediate response to loss is denial. As discussed in Chapters 3 and 5, this response is not adaptive over time, but it allows the bereaved to function immediately after the loss. The longer-term goal is to recognize and accept the loss and to return to normal functioning. The active, goal-directed behavior so commonly used successfully in adapting to other stressful situations is useless.
The central task for the bereaved, therefore, is to reconcile themselves to a situation that cannot be changed and over which they have no control. Studies on both human beings and animals indicate that the sense of control is an important variable in the psychological and physi ologic consequences of stress. Uncontrollable shock in animals has been demonstrated to be more effective than controllable shock in the promotion of gastric ulcerations, 83 immunosuppression, 55 tumor growth, 36,74 and a depression-like syndrome. 84 In animals given uncontrollable shock, Weiss et al. 84 observed similar behavior to the criteria for major depression as defined in the American Psychiatric Association's Diagnostic and Statistical Manual. Neuroendocrine changes were also correlated with both. In observing the development of gastric ulceration in rats subjected to inescapable shock, Engel 19 proposed that
to be without the information and/or the psychological behavioral resources to cope effectively with a particular life circumstance appears to be a highly significant variable. Under such conditions, the organism typically alternates between activity and inactivity, the first reflecting efforts to maintain or reestablish control, the second waiting or giving up. If its efforts succeed, or if circumstances change soon enough, illness may be averted. But the longer such cycles of struggling and inactivity go on, the greater the risk of morbidity. Several human studies indicate that it is the subjective experience, rather than the external situation, which is the best predictor of a person's reaction, be it emotional or physical.
There is physiologic evidence indicating that responses to the stresses of bereavement are not the same for all people and that individual responses are situation-specific. In comparing psychological and endocrinologic data on parents before and after loss of their children, Hofer et al. 33 emphasized that the "chronic stress mean" of a person anticipating loss applied only to that situation and could not be applied to other stressful experiences. Schleifer et al. 71 arrived at the same conclusion in studying lymphocyte activity before and after death of a spouse. Mason 51 has shown that hormonal response patterns vary, depending on the stimulus. He described several situations of "stress" (such as acute avoidance, sleep deprivation, and fasting) in which the patterns of cortisol, norepinephrine, and epinephrine are different.
Kuhn and his colleagues 44 also observed different physiologic responses, as measured by ornithine decarboxylase activity, in response to maternal separation and other types of stress in rats. Both Engel 19 and Ader and Cohen 4 noted that the relationship between "stress" and disease in animals depended on both the type of stress and the nature of the disease. Thus, it cannot be assumed that because one physiologic pattern is present in response to one type of stress, it is the prototype for all stress responses. The use of inescapable or uncontrollable punishment in animal studies appears to result in a type of stress similar to the hopelessness-helplessness syndrome described by Engel and Schmale. 20 These studies may be helpful in providing a conceptual framework from which to investigate the biology of grieving, in particular the response to unavoidable stress.
Social Relationships as Biological Regulators
Disruption of Biological Rhythms. As described earlier in this chapter and in several previous chapters, people typically have a number of symptomatic responses to bereavement. These include physiologic, behavioral, cognitive, and perceptual disturbances as well as a disorganization of ego functioning. Although the links between all these changes are not well understood, it has been hypothesized that social relationships may be an important regulator of these various responses.
Reviewing animal studies and some human studies, Hofer 32 summarized the behavioral and physiologic components of the infant separation response and called attention to the similarities between it and the human adult response to bereavement ( Table 2).
In studies on infant rats, there is a clear difference between the acute and chronic phases of the separation response; the phenomena of the slow phase remain even when the acute phase is prevented. 27-31,76 What Hofer has suggested is that the slow phase response is actually the sum of many responses to the loss of many separate aspects of the motherinfant interaction. He found that preservation of a single aspect of the mother-infant relationship could prevent a single physiologic change, without affecting any other of the physiologic responses to separation. From analytic studies on infant rats separated from their mothers, a number of "hidden regulators" have been discovered within the mother-infant relationship, which act to maintain the homeostasis of the developing child.
There is evidence of biologically powerful sensorimotor regulators even within adult social relationships, such as the menstrual synchrony that develops in young women who live together, 54 and bereavement may constitute a loss of these regulators as well as an emotional loss. The data ( Table 3) from studies of sensory deprivation, jet lag, and work shift changes can be used to emphasize the importance of unseen regulators in human behavior and to illustrate the similarities in the physiologic responses to these varied disturbances of internal equilibrium.
TABLE 3
Responses to Disrupted Regulation of Homeostasis (Human Studies).
There is an obvious similarity between these responses and the cognitive and perceptual disturbances seen in the chronic grief response, all of which contribute to the feeling of losing control. The similarities in physiologic disturbances resulting from bereavement, sensory deprivation, jet lag, and work shift change have led Hofer 32 to postulate that the regulators' function was to maintain the biological rhythms of the individual. The death of someone with whom a person has lived in close proximity involves the loss of social entraining stimuli for circadian systems, and may therefore disrupt normal biological timing. In fact, there is evidence that social interactions for human beings may be the cues regulating human biological rhythms, rather than light or temperature as in other species.
Hofer 32 suggests that these somatosensory regulators may be the precursors of psychological regulators that take over as the infant matures. He contends that disturbances of physiologic regulation may be involved even in the loss of someone who has been living far away, because internal representations of the person may have served a regulatory function; when these are altered in the psychological process of grieving, their regulatory function may be dissolved. Study of the regulatory aspects of social relationships may eventually lead to some understanding of the mechanisms by which the presence or absence of social support systems modifies physiologic responses to stress and vulnerability to disease.
Rhythms in the Endocrine and Immune Systems. It is well known that cortisol levels in plasma and urine are cyclical. There is evidence that many other hormones and biochemical systems also have cyclical patterns. 56 Catecholamine excretion displays periodicity, for example, and growth hormone secretion, although not itself rhythmic, changes in response to other variables that are in turn rhythmic. The cycles of all hormones are influenced by many environmental variables, and it seems likely that social interactions and their sudden alteration after bereavement may exert major disruptive effects.
The immune system also follows a daily rhythm. Tavadia and colleagues 79 found that the circadian rhythm followed by cortisol secretion is accompanied by changes in the number of lymphocytes and in mitogen-stimulated lymphocyte transformation. Cortisol and lymphocyte transformation were directly correlated, but both varied inversely with absolute numbers of lymphocytes. Maestroni and Pierpaoli 50 suggest that the lymphopenia seen after pinealectomy may be due to the loss of rhythmicity previously regulated by the pineal body. Others 58 have found circadian rhythms involved in immune competence, and Kort and Weijma 38 recently demonstrated that the stress of a chronic lightdark cycle shift in rats resulted in decreased lymphocyte response to a mitogen and decreased immune response as measured by popliteal lymph node assay, but not in a change in adrenocortical activity.
Grief and Depression on a Neurobiological Continuum
As discussed in the section on promising research approaches, the phenomena of bereavement mirror to a considerable extent the symptoms and signs of depressive illness and share some aspects of anxiety states. Because overt depressive illness and, to a lesser extent, severe anxiety states are often precipitated by major losses, it seems reasonable to look for connections between bereavement and these two common psychiatric disorders. Indeed, it is a plausible hypothesis that the three conditions share a common neurobiological substrate.
Klein 37 has made a strong case for this point of view, and his conceptual basis for this involves the temporal contiguity of the protest and despair phases in infantile separation responses as early "Anlagen" of the adult conditions. Klein follows Bowlby's view that these two infantile responses are deeply embedded in phylogenetic history, and hypothesizes that some individuals, even when they are adults, have low thresholds for elicitation. In some, the system may even discharge spontaneously, resulting in classical panic attacks. As Klein points out, patients with panic attacks also have an increased incidence of depressive episodes. But it was the finding that the same drug, the tricyclic antidepressant, imipramine, prevented panic attacks as well as depression that prompted Klein to develop his theory. He supposed that imipramine raised the threshold to both conditions by acting on a common neurochemical substrate.
Other findings tend to link depressive illness with bereavement. Both conditions seem to imperil the resistance of an individual to major physical challenges and have been associated with increased incidence and severity of a number of medical conditions. Second, both conditions result in increased rates of suicide. In addition, there is some limited evidence for similarity in neuroendocrine responses. Thus, in women undergoing psychotherapy for reactive depression, Sachar and his collaborators 69 noted a peak in corticosteroids when patients confronted the fact of the precipitating loss during therapy. The bereavement is often considered more painful than the chronic depression after loss, which may be defensive and help the person to avoid experiencing the loss.
Abnormal endocrine patterns are present in major affective disorders such as depression. Abnormalities in growth hormone, insulin, and thyroid-stimulating hormone are often observed in depression. 67 Depressed individuals tend to have high ACTH and cortisol levels and fail to follow normal diurnal rhythmicity; many severely depressed people fail to depress cortisol levels in response to an exogenous corticosteroid, dexamethasone. However, a new study 40 from Yale of the dexamethasone suppression test in bereaved individuals who met the criteria for a diagnosis of major depression revealed the great majority as having normal adrenocortical responses—not those considered typical of depression. 40 On this dimension, then, severe grief reactions would seem to occupy a place on a neurobiologic continuum that is distinct from the one occupied by major depression.
This study notwithstanding, most of the evidence suggests that bereavement may bear more than a superficial resemblance to depressive illness and that it would be appropriate to continue to search in the bereaved for some of the biological phenomena that have been found in depressed patients and in those with panic disorders.
CONCLUSIONS
Very little is known about the biology of the varying states of grief. Clues about promising avenues for research in this field can be gleaned from the symptomatology and epidemiology of bereavement and from current knowledge about several of the body's regulatory systems. Active grieving produces a number of symptoms that suggest involvement of the respiratory, autonomic, and endocrine systems. The epidemiology of bereavement suggests that cardiovascular and immune function may be substantially altered by grief. And there are enough similarities between grief and depression to merit a comparison of neuroendocrine and other biological changes in the two conditions.
Studies are needed on basic neurophysiologic parameters, including cardiovascular, thermal, and central nervous system regulation. In particular, more information is needed on the long-term effects of bereavement: how do physiologic responses change during prolonged grief, both "spontaneously" and in response to specific stimuli and to other significant life events? As more substantial baseline material is collected, possible genetic and experiential precursors should be sought to account for the variability in response encountered so often in studies of both animals and human beings. Considerable expansion of the knowledge of neurochemical changes accompanying the response to loss is required, including further investigation of neuroendocrine functions across a wider array of hormones as well as more studies of the neurotransmitter alterations that may underlie the dramatic and more subtle affective, motivational, and cognitive changes that emerge. In each of these areas, further development of remote, miniaturized blood collection devices (which would permit study of free-moving subjects, as in neurophysiologic research) would provide substantial opportunities to gather significant new data.
The preliminary data now available make it clear that traumatic loss experiences may have a long-term impact on the body's immune system. Expanding the data base on the disease susceptibility of bereaved subjects under controlled conditions and gaining further understanding of the basic immune processes that are affected will improve the chances that appropriate intervention models are developed.
The relationship between the responses to loss and the patterns observed in response to other life stresses should be established more fully. Do the patterns during bereavement parallel those in other situations in which "loss of control" is a prominent element? What types of physical and social environmental factors alter the intensity and time course of human physiologic responses? What is the meaning, in terms of basic processes and the possibilities for effective intervention and treatment strategies, of the fact that events at the behavioral and physiologic level are not always as congruent as might otherwise be expected? Research involving animal models plays a crucial role in generating new hypotheses and methods. But even more urgent is the need for prospective human studies employing the most modern biological and psychological methods and concepts.
REFERENCES
- 1.
- Ackerman, S.H., Keller, S.E., Schleifer, S.J., Shindledecker, R.D., Camerino, W.S., Hofer, M.A., Weiner, H., and Stein, M. Effect of premature weaning on lymphocyte stimulation in the rat. (Abstract.) Psychosomatic Medicine 45: 75, 1983.
- 2.
- Ader, A. (Ed.) Psychoneuroimmunology . New York: Academic Press, 1981.
- 3.
- Ader, R. Stress and illness: immune processes. Paper prepared for the Institute of Medicine, Washington, D.C., 1981.
- 4.
- Ader R., and Cohen, N. Behaviorally conditioned immunosuppression. Psychosomatic Medicine 37: 333-340, 1975. [PubMed: 1162023]
- 5.
- Bartrop, R., Luckhurst, E., Lazarus, L., Kiloh, L.G., and Perry, R. Depressed lymphocyte function after bereavement. Lancet 1: 834-836, 1977. [PubMed: 67339]
- 6.
- Black, S., Humphrey, J.H., and Niven, J.S.F. Inhibition of mantoux reaction by direct suggestion under hypnosis. British Medical Journal 5346: 1649-1652, 1963. [PMC free article: PMC2124383] [PubMed: 13971374]
- 7.
- Board, F., Persky, H., and Hamburg, D.A. Psychological stress and endocrine functions: blood levels of adrenocortical and thyroid hormones in acutely disturbed patients. Psychosomatic Medicine 18: 324-333, 1956. [PubMed: 13350458]
- 8.
- Breese, G.R., Smith, R.D., Mueller, R.A., Howard, J.L., Prangle, A.J., Lipton, M.A., Young, L.D., McKinney, W.T., and Lewis, J.K. Induction of adrenal catecholamine synthesizing enzymes following mother-infant separation. Nature New Biology 246: 94-96, 1973. [PubMed: 4148539]
- 9.
- Cannon, W.B. Bodily Changes in Pain, Hunger, Fear and Rage (2nd edition). New York: Appleton, 1929.
- 10.
- Cannon, W.B. Stresses and strains of homeostasis. American Journal of Medical Science 189: 1-14, 1935.
- 11.
- Case, R.M., and Sachar, E.J. Psychoendocrinology. In: Textbook of Endocrinology (6th edition) (Williams, R.H., editor. , ed.). Philadelphia: W.B. Saunders, 1981.
- 12.
- Chambers, W.N., and Reiser, M.F. Emotional stress in the precipitation of congestive heart failure. Psychosomatic Medicine 15: 38-60, 1953. [PubMed: 13027505]
- 13.
- Clayton, P.J. Mortality and morbidity in the first year of widowhood. Archives of General Psychiatry 30: 747-750, 1974. [PubMed: 4832181]
- 14.
- Coe, C.L., Mendoza, S.P., Smotherman, W.P., and Levine, S. Mother-infant attachment in the squirrel monkey: adrenal responses to separation. Behavioral Biology 22: 256-263, 1978. [PubMed: 415729]
- 15.
- Coe, C.L., Wiener, S.G., and Levine, S. Psychoendocrine responses of mother and infant monkeys to disturbance and separation. In: Symbiosis in Parent-Offspring Interaction (Rosenblum, L.A., editor; , and Moltz, H., editor. , eds.). New York: Plenum, 1983.
- 16.
- Craddock, G.C. Corticosteroid-induced lymphopenia, immunosuppression, and body defense. Annals of Internal Medicine 88: 564-566, 1978. [PubMed: 345907]
- 17.
- Elliott, G., editor; , and Eisdorfer, C., editor. (Eds.) Stress and Human Health: A Study by the Institute of Medicine . New York: Springer, 1982.
- 18.
- Engel, G. Sudden and rapid death during psychological stress. Annals of Internal Medicine 74: 771-782, 1971. [PubMed: 5559442]
- 19.
- Engel, G. Memorial Lecture: the psychosomatic approach to individual susceptibility to disease. Gastroenterology 67: 1085-1093, 1974. [PubMed: 4473394]
- 20.
- Engel, G., and Schmale, A. Conservation-withdrawal: a primary regulatory process for organismic homeostasis. Physiology, Emotion and Psychosomatic Illness (Ciba Foundation Symposium No. 8). New York: Elsevier, 1972. [PubMed: 4144967]
- 21.
- von Euler, U.S., Gemzell, C.A., Levi, L., and Strom, G. Cortical and medullary adrenal activity in emotional stress. Acta Endocrinologica 30: 567-573, 1959. [PubMed: 13636773]
- 22.
- Franksson, C., and Gemzell, C. Adrenocortical activity in preoperative period. Journal of Clinical Endocrinology and Metabolism 15: 1069-1072, 1955. [PubMed: 13263399]
- 23.
- Gala, R.R., and Haisenleder, D.J. Stress-induced decrease of the afternoon prolactin surge. Life Science 31: 875-879, 1982. [PubMed: 6891013]
- 24.
- Hattori, T., Hamai, Y., Ikeda, T., Takiyama, W., Hirai, T., and Miyoshi, Y. Inhibitory effects of immunopotentiators on the enhancement of lung metastases induced by operative stress in rats. Gan 73: 132-135, 1982. [PubMed: 7117743]
- 25.
- Hennessy, M.B., Kaplan, J.N., Mendoza, S.P., Lowe, E.L., and Levine, S. Separation distress and attachment in surrogate-reared squirrel monkeys. Physiological Behavior 23: 1017-1023, 1979. [PubMed: 120532]
- 26.
- Hofer, M.A. The role of nutrition in the physiological and behavioral effects of early maternal separation in infant rats. Psychosomatic Medicine 35: 350-359, 1973. [PubMed: 4198119]
- 27.
- Hofer, M.A. The effects of brief maternal separations on behavior and heart rate of 2 week old rat pups. Physiological Behavior 10: 423-427, 1973. [PubMed: 4708508]
- 28.
- Hofer, M.A. Survival and recovery of physiologic functions after early maternal separation in infant rats. Psychosomatic Medicine 15: 475-480, 1975. [PubMed: 1221454]
- 29.
- Hofer, M.A. Studies on how early maternal separation produces behavioral change in young rats. Psychosomatic Medicine 37: 245-246, 1975. [PubMed: 1178795]
- 30.
- Hofer, M.A. The organization of sleep and wakefulness after maternal separation in young rats. Developmental Psychobiology 9: 189-206, 1976. [PubMed: 964463]
- 31.
- Hofer, M.A. On the relationship between attachment and separation processes in infancy. In: Emotion: Theory Research and Experience , Vol. 2; Early Development (Plutchik, R., editor. , ed.). New York: Academic Press, 1983.
- 32.
- Hofer, M.A. Relationships as regulators: a psychobiological perspective on bereavement. Psychosomatic Medicine (in press), 1984. [PubMed: 6739679]
- 33.
- Hofer, M.A., Wolff, C.T., Friedman, S.B., and Mason, J.W. A psychoendocrine study of bereavement. Part I: 17-hydroxycorticosteroid excretion rates of parents following death of their children from leukemia. Psychosomatic Medicine 34: 481-491, 1972. [PubMed: 4644657]
- 34.
- Hofer, M.A., Wolff, C.T., and Mason, J.W. A psychoendocrine study of bereavement. Part II: observations on the process of mourning in relation to adrenocortical function. Psychosomatic Medicine 34: 492-504, 1972. [PubMed: 4644658]
- 35.
- Jacobs, S., Mason, J.W., Kosten, T.R., Ostfeld, A.M., Kasl, S.V., Atkins, S.R., Gardner, C.W., and Schreiber, S.J. Bereavement, psychological distress, ego defenses, and adrenocortical function. Paper presented at American Psychosomatic Society Meeting, March 1984.
- 36.
- Keller, S.E., Weiss, J.M., Schleifer, S.J., Miller, N.E., and Stein, M. Suppression of immunity by stress: effect of a graded series of stressors on lymphocyte stimulation in the rat. Science 213: 1397-1400, 1981. [PubMed: 6973822]
- 37.
- Klein, D.F. Anxiety reconceptualized. In: Anxiety: New Research and Changing Concepts (Klein, D.F., editor; , and Rabkin, J., editor. , eds.). New York: Raven Press, 1981.
- 38.
- Kort, W.J., and Weijma, J.M. Effect of chronic light-dark shift stress on the immune response of the rat. Physiological Behavior 29: 1083-1087, 1982. [PubMed: 7163387]
- 39.
- Kosten, T.R., Jacobs, S., and Mason, J.W. Psychological correlates of growth hormone response to stress. (Abstract.) Psychosomatic Medicine 45: 82, 1983.
- 40.
- Kosten, T.R., Jacobs, S., and Mason, J. The dexamethasone suppression test during bereavement. Journal of Nervous and Mental Diseases (in press). [PubMed: 6544808]
- 41.
- Kosten, T.R., Jacobs, S., Mason, J., and Wahby, V. Growth hormone response during the stress of bereavement. Paper presented at American Psychosomatic Society Meeting, March 1984.
- 42.
- Kraemer, G., and McKinney, W. Interaction of pharmacological agents which alter biogenic amine metabolism and depression. Journal of Affective Disorders 1: 33-54, 1979. [PubMed: 45183]
- 43.
- Kreuz, L., Rose, R., and Jennings, J. Suppression of plasma testosterone levels and psychological stress. Archives of General Psychiatry 25: 479-482, 1972. [PubMed: 5019887]
- 44.
- Kuhn, C.M., Grignolo, A., Johnson, M., and Schanberg, S.M. Heterogeneity in the response pattern of organ systems to stress: mediators and mechanisms. Psychopharmacological Bulletin 18: 96-99, 1982.
- 45.
- Lader, M.H. Palmar skin conductance measures in anxiety and phobic states. Journal of Psychosomatic Research 11: 271-281, 1967. [PubMed: 6076920]
- 46.
- Laudenslager, M., Reite, M., and Harbeck, M. Suppressed immune response in infant monkeys associated with maternal separation. Behavioral and Neural Biology 36: 40-48, 1982. [PubMed: 7168729]
- 47.
- Levine, S., and Coe, C.L. Recent advances in psychophysiology: endocrine regulation. In: Handbook of Psychosomatic Medicine (Cheren, S., editor. , ed.). New York: State University of New York Press; (in press).
- 48.
- Lindemann, E. The symptomatology and management of acute grief. American Journal of Psychiatry 101: 141-148, 1944.
- 49.
- Lown, B., Venner, R.L., and Corbalan, R. Psychologic stress and threshold for repetitive ventricular response. Science 182: 834-836, 1973. [PubMed: 4795748]
- 50.
- Maestroni, G.J.M., and Pierpaoli, W. Pharmacologic control of the hormonally mediated immune response. In: Psychoneuroimmunology (Ader, A., editor. , ed. . New York: Academic Press, 1981.
- 51.
- Mason, J.W. Emotion as reflected in patterns of endocrine integration. In: Emotions—Their Parameters and Measurement (Levi, L., editor. , ed.). New York: Raven Press, 1975.
- 52.
- Mason, J.W. Bereavement: endocrine aspects. Paper presented at American Psychosomatic Society Meeting, March 1983.
- 53.
- Mason, J.W., Sachar, E.J., Fishman, J.R., Hamburg, D.A., and Handlon, J.H. Corticosteroid responses to hospital admission. Archives of General Psychiatry , 13: 1-8, 1965. [PubMed: 14303983]
- 54.
- 55.
- Monjan, A.A. Stress and immunologic competence: studies in animals. In: Psychoneuroimmunology (Ader, A., editor. , ed.). New York: Academic Press, 1981.
- 56.
- Moore-Ede, M.C., Sulzman, F.M., and Fuller, C.A. The Clocks that Time Us . Cambridge, MA: Harvard University Press, 1982.
- 57.
- Newberry, B.H. Effects of presumably stressful stimulation (PSS} on the development of animal tumors: some issues. In: Perspectives on Behavioral Medicine (Weiss, S.M., editor; , Herd, J.A., editor; , and Fox, B.H., editor. , eds.). New York: Academic Press, 1981.
- 58.
- Palmblad, J. Stress and immunologic competence: studies in man. In: Psychoneuroimmunology (Ader, A., editor. , ed.). New York: Academic Press, 1981.
- 59.
- Parkes, C.M. Bereavement: Studies of Grief in Adult Life. London: Tavistock, 1972.
- 60.
- Reich, P. Clinical observations on the psychobiology of life-threatening arrhythmias. In: Biobehavioral Factors in Sudden Cardiac Death (Solomon, F., editor; , Parron, D.L., editor; , and Dews, P.B., editor. , eds.). Washington, D.C.: National Academy Press, 1982.
- 61.
- Reich, P., DeSilva, R.A., Lown, B., and Murawski, B.J. Acute psychological disturbance preceding life-threatening ventricular arrhythmias. Journal of the American Medical Association 246: 233-235, 1981. [PubMed: 7241762]
- 62.
- Reite, M., Kaufman, K., Pauley, J.D., and Stynes, A.J. Depression in infant monkeys: physiological correlates. Psychosomatic Medicine 36: 363-367, 1974. [PubMed: 4369909]
- 63.
- Reite, M., and Short, R.A. Nocturnal sleep in separated monkey infants. Archives of General Psychiatry 35: 1247-1253, 1973. [PubMed: 211985]
- 64.
- Riley, V. Psychoneuroendocrine influences on immuno-competence and neoplasia. Science 212: 1100-1109, 1981. [PubMed: 7233204]
- 65.
- Riley, V., Fitzmaurice, M.A., and Spackman, D.H. Psychoneuroimmunologic factors in neoplasia: studies in animals. In: Psychoneuroimmunology (Ader, A., editor. , ed.). New York: Academic Press, 1981.
- 66.
- Rose, R.M., Poe, R.O., and Mason, J.W. Psychological state and body size as determinants of 17-OHCS excretion. Archives of Internal Medicine 121: 406-413, 1968. [PubMed: 5645717]
- 67.
- Rose, R.M., and Sachar. E. Psychoendocrinology. In: Textbook of Endocrinology (Williams, R.H., editor. , ed.) Philadelphia: W.B. Saunders, 1981.
- 68.
- Saavedra, J.M. Changes in dopamine, noradrenaline and adrenaline in specific septal and preoptic nuclei after acute immobilization stress. Neuroendocrinology 33: 396-401, 1982. [PubMed: 7145030]
- 69.
- Sachar, E.J., Mackenzie, J.M., Binstock, W.A., and Mack, J.E. Corticosteroid responses to the psychotherapy of reactive depressions. Psychosomatic Medicine 30: 23-44, 1968. [PubMed: 5640284]
- 70.
- Schiffman, P.L., Westlake, R.E., Santiago, T.V., and Edelman, N.H. Ventilation control of parents of victims of Sudden Infant Death Syndrome. New England Journal of Medicine 302: 486-491, 1980. [PubMed: 7351974]
- 71.
- Schleifer, S.J., Keller, S.E., Camerino, M., Thornton, J.C., and Stein, M. Suppression of lymphocyte stimulation following bereavement. Journal of the American Medical Association 250: 374-399, 1983. [PubMed: 6854901]
- 72.
- Solomon, F., editor; , Parron, D.L., editor; , and Dews, P.B., editor. (Eds.) Biobehavioral Factors in Sudden Cardiac Death: Summary of an Institute of Medicine Conference . Washington, D.C.: National Academy Press, 1982.
- 73.
- Solomon, G.F., and Amkraut, A.A. Emotions, stress, and immunity. In: Frontiers of Radiation in Therapeutic Oncology , Vol. 7 (Vaeth, J.M., editor. , ed.). Baltimore: University Park Press, 1972.
- 74.
- Sowers, J.R., Raj, R.P., Hershman, J.M., Carlson, H.E., and McCallum, R.W. The effect of stressful diagnostic studies and surgery on anterior pituitary hormone release in man. Acta Endocrinologica 86: 25-32, 1977. [PubMed: 578611]
- 75.
- Starkman, M.N., Schteingart, D.E., and Schork, M.A. Depressed mood and other psychiatric manifestations of Cushing's syndrome: relationship to hormone levels. Psychosomatic Medicine 43: 3-18, 1981. [PubMed: 6259680]
- 76.
- Stone, E., Bonnet, K., and Hofer, M.A. Survival and development of maternally deprived rats: role of body temperature. Psychosomatic Medicine 38: 242-249, 1976. [PubMed: 940904]
- 77.
- Suomi, S.J., Seaman, S.F., Lewis, J.K., Delizio, R.D., and McKinney, W.T., Jr. Effects of imipramine treatment of separation-induced social disorders in rhesus monkeys. Archives of General Psychiatry 35: 321-327, 1978. [PubMed: 103510]
- 78.
- Sutton, J.R., and Casey, J.H. The adrenocortical response to competitive athletics in veteran athletes. Journal of Clinical Endocrinology and Metabolism 40: 135-138, 1975. [PubMed: 1112872]
- 79.
- Tavadia, H.B., Fleming, K.A., Hume, P.D., and Simpson, H.W. Circadian rhythmicity of human plasma cortisol and PHA-induced lymphocyte transformation. Clinical and Experimental Immunology 22: 190-193, 1975. [PMC free article: PMC1538331] [PubMed: 1212815]
- 80.
- Verrier, R.L., and Lown, B. Autonomic nervous system and malignant cardiac arrhythmias. In: Brain, Behavior and Bodily Disease (Weiner, H., editor; , Hofer, M.A., editor; , and Stunkard, A.J., editor. , eds.). New York: Raven Press, 1981.
- 81.
- Vogt, J.L., and Hennessy, M.B. Infant separation in monkeys: studies on social figures other than the mother. In: Child Nurturance: Studies of Development in Nonhuman Primates (Fitzgerald, H.E., editor; , Mullins, J.A., editor; , and Gage, P., editor. , eds.). New York: Plenum, 1982.
- 82.
- Weiner, H., Singer, M.T., and Reiser, M.F. Cardiovascular responses and their psychological correlates: a study in healthy young adults and patients with peptic ulcer and hypertension. Psychosomatic Medicine 24: 477-498, 1962. [PubMed: 13999596]
- 83.
- Weiss, J.M. The effects of coping responses on stress. Journal of Comparative Physiology and Psychology 65: 251-260, 1968. [PubMed: 5668311]
- 84.
- Weiss, J.M., Bailey, W.H., Goodman, P.A., Hoffman, L.J., Ambrose, M.J., Salman, S., and Charry, J.M. A model for neurochemical study of depression. In: Behavioral Models and the Analysis of Drug Action (Levy, A., editor; , and Spiegelstein, M.Y., editor. , eds.). Amsterdam: Elsevier, 1982.
- 85.
- Wiener, S., Johnson, D., and Levine, S. Behavioral and physiological response of infant squirrel monkeys following weaning. Paper presented at American Society of Primatologists Meeting, East Lansing, Michigan, 1983.
- 86.
- Wolff, C.T., Hofer, M.A., Friedman, S.B., and Mason, J.W. Relationships of psychological defenses and mean urinary 17-hydroxycorticosteroid excretion rates in the parents of children with leukemia. Parts I and II. Psychosomatic Medicine 26: 576-609, 1964. [PubMed: 14219239]
Footnotes
This chapter is based on material prepared by committee members Jules Hirsch, M.D., Myron Hofer, M.D., and Jimmie Holland, M.D., in collaboration with Fredric Solomon, M.D. It draws upon a background paper by Leonard Rosenblum, Ph.D., Director, Primate Behavior Laboratory, Downstate Medical Center, Brooklyn, New York. Additional assistance and background materials were provided by Elizabeth Guilfoyle, medical student, New York University School of Medicine.
- Toward a Biology of Grieving - BereavementToward a Biology of Grieving - Bereavement
Your browsing activity is empty.
Activity recording is turned off.
See more...