By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Copyright © 2000, Geoffrey M Cooper.
Bookshelf ID: NBK9961

An official website of the United States government

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Cooper GM. The Cell: A Molecular Approach. 2nd edition. Sunderland (MA): Sinauer Associates; 2000.

Actin filaments, usually in association with myosin, are responsible for many types of cell movements. Myosin is the prototype of a molecular motor—a protein that converts chemical energy in the form of ATP to mechanical energy, thus generating force and movement. The most striking variety of such movement is muscle contraction, which has provided the model for understanding actin-myosin interactions and the motor activity of myosin molecules. However, interactions of actin and myosin are responsible not only for muscle contraction but also for a variety of movements of nonmuscle cells, including cell division, so these interactions play a central role in cell biology. Moreover, the actin cytoskeleton is responsible for the crawling movements of cells across a surface, which appear to be driven directly by actin polymerization as well as actin-myosin interactions.
Muscle cells are highly specialized for a single task, contraction, and it is this specialization in structure and function that has made muscle the prototype for studying movement at the cellular and molecular levels. There are three distinct types of muscle cells in vertebrates: skeletal muscle, which is responsible for all voluntary movements; cardiac muscle, which pumps blood from the heart; and smooth muscle, which is responsible for involuntary movements of organs such as the stomach, intestine, uterus, and blood vessels. In both skeletal and cardiac muscle, the contractile elements of the cytoskeleton are present in highly organized arrays that give rise to characteristic patterns of cross-striations. It is the characterization of these structures in skeletal muscle that has led to our current understanding of muscle contraction, and other actin-based cell movements, at the molecular level.
Skeletal muscles are bundles of muscle fibers, which are single large cells (approximately 50 μm in diameter and up to several centimeters in length) formed by the fusion of many individual cells during development (Figure 11.18). Most of the cytoplasm consists of myofibrils, which are cylindrical bundles of two types of filaments: thick filaments of myosin (about 15 nm in diameter) and thin filaments of actin (about 7 nm in diameter). Each myofibril is organized as a chain of contractile units called sarcomeres, which are responsible for the striated appearance of skeletal and cardiac muscle.

Structure of muscle cells. Muscles are composed of bundles of single large cells (called muscle fibers) that form by cell fusion and contain multiple nuclei. Each muscle fiber contains many myofibrils, which are bundles of actin and myosin filaments organized (more...)
The sarcomeres (which are approximately 2.3 μm long) consist of several distinct regions, discernible by electron microscopy, which provided critical insights into the mechanism of muscle contraction (Figure 11.19). The ends of each sarcomere are defined by the Z disc. Within each sarcomere, dark bands (called A bands because they are anisotropic when viewed with polarized light) alternate with light bands (called I bands for isotropic). These bands correspond to the presence or absence of myosin filaments. The I bands contain only thin (actin) filaments, whereas the A bands contain thick (myosin) filaments. The myosin and actin filaments overlap in peripheral regions of the A band, whereas a middle region (called the H zone) contains only myosin. The actin filaments are attached at their plus ends to the Z disc, which includes the crosslinking protein α-actinin. The myosin filaments are anchored at the M line in the middle of the sarcomere.

Structure of the sarcomere. (A) Electron micrograph of a sarcomere. (B) Diagram showing the organization of actin (thin) and myosin (thick) filaments in the indicated regions. (A, Frank A. Pepe/Biological Photo Service.)
Two additional proteins (titin and nebulin) also contribute to sarcomere structure and stability (Figure 11.20). Titin is an extremely large protein (3000 kd), and single titin molecules extend from the M line to the Z disc. These long molecules of titin are thought to act like springs that keep the myosin filaments centered in the sarcomere and maintain the resting tension that allows a muscle to snap back if overextended. Nebulin filaments are associated with actin and are thought to regulate the assembly of actin filaments by acting as rulers that determine their length.

Titin and nebulin. Molecules of titin extend from the Z disc to the M line and act as springs to keep myosin filaments centered in the sarcomere. Molecules of nebulin extend from the Z disc and are thought to determine the length of associated actin filaments. (more...)
The basis for understanding muscle contraction is the sliding filament model, first proposed in 1954 both by Andrew Huxley and Ralph Niedergerke and by Hugh Huxley and Jean Hanson (Figure 11.21). During muscle contraction, each sarcomere shortens, bringing the Z discs closer together. There is no change in the width of the A band, but both the I bands and the H zone almost completely disappear. These changes are explained by the actin and myosin filaments sliding past one another, so that the actin filaments move into the A band and H zone. Muscle contraction thus results from an interaction between the actin and myosin filaments that generates their movement relative to one another. The molecular basis for this interaction is the binding of myosin to actin filaments, allowing myosin to function as a motor that drives filament sliding.
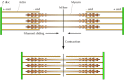
Sliding-filament model of muscle contraction. The actin filaments slide past the myosin filaments toward the middle of the sarcomere. The result is shortening of the sarcomere without any change in filament length.
The type of myosin present in muscle (myosin II) is a very large protein (about 500 kd) consisting of two identical heavy chains (about 200 kd each) and two pairs of light chains (about 20 kd each) (Figure 11.22). Each heavy chain consists of a globular head region and a long α-helical tail. The α-helical tails of two heavy chains twist around each other in a coiled-coil structure to form a dimer, and two light chains associate with the neck of each head region to form the complete myosin II molecule.

Myosin II. The myosin II molecule consists of two heavy chains and two pairs of light chains (called the essential and regulatory light chains). The heavy chains have globular head regions and long α-helical tails, which coil around each other (more...)
The thick filaments of muscle consist of several hundred myosin molecules, associated in a parallel staggered array by interactions between their tails (Figure 11.23). The globular heads of myosin bind actin, forming cross-bridges between the thick and thin filaments. It is important to note that the orientation of myosin molecules in the thick filaments reverses at the M line of the sarcomere. The polarity of actin filaments (which are attached to Z discs at their plus ends) similarly reverses at the M line, so the relative orientation of myosin and actin filaments is the same on both halves of the sarcomere. As discussed later, the motor activity of myosin moves its head groups along the actin filament in the direction of the plus end. This movement slides the actin filaments from both sides of the sarcomere toward the M line, shortening the sarcomere and resulting in muscle contraction.

Organization of myosin thick filaments. Thick filaments are formed by the association of several hundred myosin II molecules in a staggered array. The globular heads of myosin bind actin, forming cross-bridges between the myosin and actin filaments. The (more...)
In addition to binding actin, the myosin heads bind and hydrolyze ATP, which provides the energy to drive filament sliding. This translation of chemical energy to movement is mediated by changes in the shape of myosin resulting from ATP binding. The generally accepted model (the swinging-cross-bridge model) is that ATP hydrolysis drives repeated cycles of interaction between myosin heads and actin. During each cycle, conformational changes in myosin result in the movement of myosin heads along actin filaments.
Although the molecular mechanisms are still not fully understood, a plausible working model for myosin function has been derived both from in vitro studies of myosin movement along actin filaments (a system developed by James Spudich and Michael Sheetz) and from determination of the three-dimensional structure of myosin by Ivan Rayment and his colleagues (Figure 11.24). The cycle starts with myosin (in the absence of ATP) tightly bound to actin. ATP binding dissociates the myosin-actin complex and the hydrolysis of ATP then induces a conformational change in myosin. This change affects the neck region of myosin that binds the light chains (see Figure 11.22), which acts as a lever arm to displace the myosin head by about 5 nm. The products of hydrolysis (ADP and Pi) remain bound to the myosin head, which is said to be in the “cocked” position. The myosin head then rebinds at a new position on the actin filament, resulting in the release of ADP and Pi and triggering the “power stroke,” in which the myosin head returns to its initial conformation, thereby sliding the actin filament toward the M line of the sarcomere.
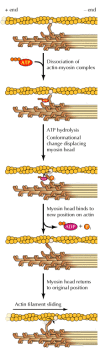
Model for myosin action. The binding of ATP dissociates myosin from actin. ATP hydrolysis then induces a conformational change that displaces the myosin head group. This is followed by binding of the myosin head to a new position on the actin filament (more...)
The contraction of skeletal muscle is triggered by nerve impulses, which stimulate the release of Ca2+ from the sarcoplasmic reticulum—a specialized network of internal membranes, similar to the endoplasmic reticulum, that stores high concentrations of Ca2+ ions. The release of Ca2+ from the sarcoplasmic reticulum increases the concentration of Ca2+ in the cytosol from approximately 10-7 to 10-5 M. The increased Ca2+ concentration signals muscle contraction via the action of two accessory proteins bound to the actin filaments: tropomyosin and troponin (Figure 11.25). Tropomyosin is a fibrous protein that binds lengthwise along the groove of actin filaments. In striated muscle, each tropomyosin molecule is bound to troponin, which is a complex of three polypeptides: troponin C (Ca2+-binding), troponin I (inhibitory), and troponin T (tropomyosin-binding). When the concentration of Ca2+ is low, the complex of the troponins with tropomyosin blocks the interaction of actin and myosin, so the muscle does not contract. At high concentrations, Ca2+ binding to troponin C shifts the position of the complex, relieving this inhibition and allowing contraction to proceed.
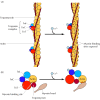
Association of tropomyosin and troponins with actin filaments. (A) Tropomyosin binds lengthwise along actin filaments and, in striated muscle, is associated with a complex of three troponins: troponin I (TnI), troponin C (TnC), and troponin T (TnT). In (more...)
Contractile assemblies of actin and myosin, resembling small-scale versions of muscle fibers, are present also in nonmuscle cells. As in muscle, the actin filaments in these contractile assemblies are interdigitated with bipolar filaments of myosin II, consisting of 15 to 20 myosin II molecules, which produce contraction by sliding the actin filaments relative to one another (Figure 11.26). The actin filaments in contractile bundles in nonmuscle cells are also associated with tropomyosin, which facilitates their interaction with myosin II, probably by competing with filamin for binding sites on actin.

Contractile assemblies in nonmuscle cells. Bipolar filaments of myosin II produce contraction by sliding actin filaments in opposite directions.
Two examples of contractile assemblies in nonmuscle cells, stress fibers and adhesion belts, were discussed earlier with respect to attachment of the actin cytoskeleton to regions of cell-substrate and cell-cell contacts (see Figures 11.13 and 11.14). The contraction of stress fibers produces tension across the cell, allowing the cell to pull on a substrate (e.g., the extracellular matrix) to which it is anchored. The contraction of adhesion belts alters the shape of epithelial cell sheets: a process that is particularly important during embryonic development, when sheets of epithelial cells fold into structures such as tubes.
The most dramatic example of actin-myosin contraction in nonmuscle cells, however, is provided by cytokinesis—the division of a cell into two following mitosis (Figure 11.27). Toward the end of mitosis in animal cells, a contractile ring consisting of actin filaments and myosin II assembles just underneath the plasma membrane. Its contraction pulls the plasma membrane progressively inward, constricting the center of the cell and pinching it in two. Interestingly, the thickness of the contractile ring remains constant as it contracts, implying that actin filaments disassemble as contraction proceeds. The ring then disperses completely following cell division.

Cytokinesis. Following completion of mitosis (nuclear division), a contractile ring consisting of actin filaments and myosin II divides the cell in two.
The regulation of actin-myosin contraction in striated muscle, discussed earlier, is mediated by the binding of Ca2+ to troponin. In nonmuscle cells and in smooth muscle, however, contraction is regulated primarily by phosphorylation of one of the myosin light chains, called the regulatory light chain (Figure 11.28). Phosphorylation of the regulatory light chain in these cells has at least two effects: It promotes the assembly of myosin into filaments, and it increases myosin catalytic activity, enabling contraction to proceed. The enzyme that catalyzes this phosphorylation, called myosin light-chain kinase, is itself regulated by association with the Ca2+-binding protein calmodulin. Increases in cytosolic Ca2+ promote the binding of calmodulin to the kinase, resulting in phosphorylation of the myosin regulatory light chain. Increases in cytosolic Ca2+ are thus responsible, albeit indirectly, for activating myosin in smooth muscle and nonmuscle cells, as well as in striated muscle.
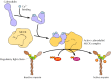
Regulation of myosin by phosphorylation. Ca2+ binds to calmodulin, which in turn binds to myosin light-chain kinase (MLCK). The active calmodulin-MLCK complex then phosphorylates the myosin II regulatory light chain, converting myosin from an inactive (more...)
In addition to myosin II (“conventional” two-headed myosin), several other types of myosin are found in nonmuscle cells. In contrast to myosin II, these “unconventional” myosins do not form filaments and therefore are not involved in contraction. They may, however, be involved in a variety of other kinds of cell movements, such as the transport of membrane vesicles and organelles along actin filaments, phagocytosis, and extension of pseudopods in amoebae (see Figure 11.17).
The best-studied of these unconventional myosins are members of the myosin I family (Figure 11.29). The myosin I proteins contain a globular head group that acts as a molecular motor, like that of myosin II. However, members of the myosin I family are much smaller molecules (about 110 kd in mammalian cells) that lack the long tail of myosin II and do not form dimers. Their tails can instead bind to other structures, such as membrane vesicles or organelles. The movement of myosin I along an actin filament can then transport its attached cargo. One function of myosin I, discussed earlier, is to form the lateral arms that link actin bundles to the plasma membrane of intestinal microvilli (see Figure 11.16). In these structures, the motor activity of myosin I may move the plasma membrane along the actin bundles, toward the tip of the microvillus. Additional functions of myosin I may be in the transport of vesicles and organelles along actin filaments and in movement of the plasma membrane during phagocytosis and pseudopod extension.
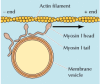
Myosin I. Myosin I contains a head group similar to myosin II, but it has a comparatively short tail and does not form dimers or filaments. Although it cannot induce contraction, myosin I can move along actin filaments (toward the plus end), carrying (more...)
In addition to myosins I and II, at least 12 other classes of unconventional myosins (III through XIV) have been identified. Some of these unconventional myosins are two-headed like myosin II, whereas others are one-headed like myosin I. The functions of most of these unconventional myosins remain to be determined, but some have been clearly shown to play important roles in organelle movement (myosins V and VI) and in sensory functions such as vision (myosin III) and hearing (myosins VI and VII).
The crawling movements of cells across a surface represent a basic form of cell locomotion, employed by a wide variety of different kinds of cells. Examples include the movements of amoebas, the migration of embryonic cells during development, the invasion of tissues by white blood cells to fight infection, the migration of cells involved in wound healing, and the spread of cancer cells during the metastasis of malignant tumors. Similar types of movement are also responsible for phagocytosis and for the extension of nerve cell processes during development of the nervous system. All of these movements are based on the dynamic properties of the actin cytoskeleton, although the detailed mechanisms involved remain to be fully understood.
Cell crawling involves a coordinated cycle of movements, which can be viewed in three stages. First, protrusions such as pseudopodia, lamellipodia, or microspikes (see Figure 11.17) must be extended from the leading edge of the cell (Figure 11.30). Second, these extensions must attach to the substratum across which the cell is migrating. Finally, the trailing edge of the cell must dissociate from the substratum and retract into the cell body.

Cell crawling. The crawling movements of cells across a surface can be viewed as three stages of coordinated movements: (1) extension of the leading edge, (2) attachment of the leading edge to the substratum, and (3) retraction of the rear of the cell (more...)
A variety of experiments indicate that extension of the leading edge involves the polymerization and crosslinking of actin filaments. For example, inhibition of actin polymerization (e.g., by treatment with cytochalasin) blocks the formation of cell surface protrusions. The regulated turnover of actin filaments, as illustrated in Figure 11.5, leads to the extension of processes such as filopodia and lamellipodia at the leading edge of the cell, and both cofilin and Arp2/3 proteins appear to be involved in this process. Unconventional myosins may also participate in the extension of processes at the leading edge: Myosin I is required for pseudopod extension in the amoeba Dictyostelium and Myosin V for extension of filopodia in neurons.
Following their extension, protrusions from the leading edge must attach to the substratum in order to function in cell locomotion. For slow-moving cells, such as fibroblasts, attachment involves the formation of focal adhesions (see Figure 11.13). Cells moving more rapidly, such as amoebas or white blood cells, form more diffuse contacts with the substratum, the molecular composition of which is not known.
The third stage of cell crawling, retraction of the trailing edge, is the least understood. The attachments of the trailing edge to the substratum are broken, and the rear of the cell recoils into the cell body. The process appears to require the development of tension between the front and rear of the cell, generating contractile force that eventually pulls the rear of the cell forward. This aspect of cell locomotion is impaired in mutants of Dictyostelium lacking myosin II, consistent with a role for myosin II in contracting the actin cortex and generating the force required for retraction of the trailing edge.
By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Your browsing activity is empty.
Activity recording is turned off.
See more...