By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Copyright © 2000, Geoffrey M Cooper.
Bookshelf ID: NBK9851

An official website of the United States government

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Cooper GM. The Cell: A Molecular Approach. 2nd edition. Sunderland (MA): Sinauer Associates; 2000.

Direct interactions between cells, as well as between cells and the extracellular matrix, are critical to the development and function of multicellular organisms. Some cell-cell interactions are transient, such as the interactions between cells of the immune system and the interactions that direct white blood cells to sites of tissue inflammation. In other cases, stable cell-cell junctions play a key role in the organization of cells in tissues. For example, several different types of stable cell-cell junctions are critical to the maintenance and function of epithelial cell sheets. Plant cells also associate with their neighbors not only by interactions between their cell walls, but also by specialized junctions between their plasma membranes.
Cell-cell adhesion is a selective process, such that cells adhere only to other cells of specific types. This selectivity was first demonstrated in classical studies of embryo development, which showed that cells from one tissue (e.g., liver) specifically adhere to cells of the same tissue rather than to cells of a different tissue (e.g., brain). Such selective cell-cell adhesion is mediated by transmembrane proteins called cell adhesion molecules, which can be divided into four major groups: the selectins, the integrins, the immunoglobulin (Ig) superfamily (so named because they contain structural domains similar to immunoglobulins), and the cadherins (Table 12.3). Cell adhesion mediated by the selectins, integrins, and cadherins requires Ca2+ or Mg2+, so many adhesive interactions between cells are Ca2+- or Mg2+-dependent.
Cell Adhesion Molecules.
The selectins mediate transient interactions between leukocytes and endothelial cells or blood platelets. There are three members of the selectin family: L-selectin, which is expressed on leukocytes; E-selectin, which is expressed on endothelial cells; and P-selectin, which is expressed on platelets. As discussed earlier in this chapter, the selectins recognize cell surface carbohydrates (see Figure 12.14). One of their critical roles is to initiate the interactions between leukocytes and endothelial cells during the migration of leukocytes from the circulation to sites of tissue inflammation (Figure 12.62). The selectins mediate the initial adhesion of leukocytes to endothelial cells. This is followed by the formation of more stable adhesions, in which integrins on the surface of leukocytes bind to intercellular adhesion molecules (ICAMs), which are members of the Ig superfamily expressed on the surface of endothelial cells. The firmly attached leukocytes are then able to penetrate the walls of capillaries and enter the underlying tissue by migrating between endothelial cells.
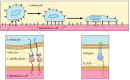
Adhesion between leukocytes and endothelial cells. Leukocytes leave the circulation at sites of tissue inflammation by interacting with the endothelial cells of capillary walls. The first step in this interaction is the binding of leukocyte selectins (more...)
The binding of ICAMs to integrins is an example of a heterophilic interaction, in which an adhesion molecule on the surface of one cell (e.g., an ICAM) recognizes a different molecule on the surface of another cell (e.g., an integrin). Other members of the Ig superfamily mediate homophilic interactions, in which an adhesion molecule on the surface of one cell binds to the same molecule on the surface of another cell. Such homophilic binding leads to selective adhesion between cells of the same type. For example, nerve cell adhesion molecules (N-CAMs) are members of the Ig superfamily expressed on nerve cells, and homophilic binding between N-CAMs contributes to the formation of selective associations between nerve cells during development. There are more than 100 members of the Ig superfamily, which mediate a variety of cell-cell interactions.
The fourth group of cell adhesion molecules, the cadherins, also display homophilic binding specificities. They are not only involved in selective adhesion between embryonic cells but are also primarily responsible for the formation of stable junctions between cells in tissues. For example, E-cadherin is expressed on epithelial cells, so homophilic interactions between E-cadherins lead to the selective adhesion of epithelial cells to one another. It is noteworthy that loss of E-cadherin can lead to the development of cancers arising from epithelial cells, illustrating the importance of cell-cell interactions in controlling cell behavior. Different members of the cadherin family, such as N-cadherin (neural cadherin) and P-cadherin (placental cadherin), mediate selective adhesion of other cell types.
About twenty different classic cadherins, such as E-cadherin, have been identified. In addition, a distinct subfamily of cadherins, called protocadherins, are expressed in the central nervous system where they appear to play a role in adhesion between neurons at synapses. Intriguingly, different neurons appear to express different protocadherins, suggesting that the protocadherins may play a role in the establishment of specific connections between neurons. About 50 human protocadherin genes have been identified and shown to be organized into three gene clusters. Each cluster contains multiple exons encoding the N-terminal extracellular and transmembrane protocadherin domains, but only a single set of three exons encoding the C-terminal cytoplasmic domain (Figure 12.63). The protocadherin gene clusters thus appear to consist of a variable region, encoding multiple extracellular and transmembrane domains, linked to a constant region encoding a single cytoplasmic domain. This organization of protocadherin genes strikingly resembles that of immunoglobulin and T-cell receptor genes (see Figures 5.42 and 5.43), in which multiple variable region exons are joined to a single constant region exon. In the immunoglobulin and T-cell receptor genes, this occurs as a result of DNA rearrangements that generate diversity in the immune system. It remains to be determined whether the variable and constant regions of protocadherins are joined at the DNA or the RNA level (for example, by alternative splicing) and to what extent rearrangements of protocadherin genes might contribute to the establishment of specific synaptic connections in the brain.

Organization of protocadherin gene clusters. The human protocadherin genes are organized into three clusters. In the cluster illustrated, 15 different variable regions encoding extracellular and transmembrane domains are linked to a single constant region, (more...)
In contrast to the stable cell-matrix junctions discussed in the preceding section, the cell-cell interactions mediated by the selectins, integrins, and members of the Ig superfamily are transient adhesions in which the cytoskeletons of adjacent cells are not linked to one another. Stable adhesion junctions involving the cytoskeletons of adjacent cells are instead mediated by the cadherins. As discussed in Chapter 11, these cell-cell junctions are of two types: adherens junctions and desmosomes, in which cadherins or related proteins (desmogleins and desmocollins) are linked to actin bundles and intermediate filaments, respectively (Figure 12.64). The role of the cadherins in linking the cytoskeletons of adjacent cells is thus analogous to that of the integrins in forming stable junctions between cells and the extracellular matrix.

Stable cell-cell junctions mediated by the cadherins. Homophilic interactions between cadherins mediate two types of stable cell-cell adhesions. In adherens junctions, the cadherins are linked to bundles of actin filaments via the catenins (see Figure (more...)
In addition to the adhesion junctions mediated by the cadherins, two other types of specialized cell-cell junctions play key roles in animal tissues. Tight junctions, which are usually associated with adherens junctions and desmosomes in a junctional complex (Figure 12.65), are critically important to the function of epithelial cell sheets as barriers between fluid compartments. For example, the intestinal epithelium separates the lumen of the intestine from the underlying connective tissue, which contains blood capillaries. Tight junctions play two roles in allowing epithelia to fulfill such barrier functions. First, tight junctions form seals that prevent the free passage of molecules (including ions) between the cells of epithelial sheets. Second, tight junctions separate the apical and basolateral domains of the plasma membrane by preventing the free diffusion of lipids and membrane proteins between them. Consequently, specialized transport systems in the apical and basolateral domains are able to control the traffic of molecules between distinct extracellular compartments, such as the transport of glucose between the intestinal lumen and the blood supply (see Figure 12.32).

Tight junctions. (A) Electron micrograph of epithelial cells joined by a junctional complex, including a tight junction, an adherens junction, and a desmosome. (B) Tight junctions are formed by interactions between strands of transmembrane proteins (occludin (more...)
Tight junctions are the closest known contacts between adjacent cells. They were originally described as sites of apparent fusion between the outer leaflets of the plasma membranes, although it is now clear that the membranes do not fuse. Instead, tight junctions appear to be formed by a network of protein strands that continues around the entire circumference of the cell (see Figure 12.65). Each strand in these networks is thought to be composed of transmembrane proteins (claudins and occludin) that bind to similar proteins on adjacent cells, thereby sealing the space between their plasma membranes.
Gap junctions, which are found in most animal tissues, serve as direct connections between the cytoplasms of adjacent cells. They provide open channels through the plasma membrane, allowing ions and small molecules (less than approximately a thousand daltons) to diffuse freely between neighboring cells, but preventing the passage of proteins and nucleic acids. Consequently, gap junctions couple both the metabolic activities and the electric responses of the cells they connect. Most cells in animal tissues—including epithelial cells, endothelial cells, and the cells of cardiac and smooth muscle—communicate by gap junctions. In electrically excitable cells, such as heart muscle cells, the direct passage of ions through gap junctions couples and synchronizes the contractions of neighboring cells. Gap junctions also allow the passage of some intracellular signaling molecules, such as cAMP and Ca2+, between adjacent cells, potentially coordinating the responses of cells in tissues.
Gap junctions are constructed of transmembrane proteins called connexins (Figure 12.66). Six connexins assemble to form a cylinder with an open aqueous pore in its center. Such an assembly of connexins in the plasma membrane of one cell then aligns with the connexins of an adjacent cell, forming an open channel between the two cytoplasms. The plasma membranes of the two cells are separated by a gap corresponding to the space occupied by the connexin extracellular domains—hence the term “gap junction,” which was coined by electron microscopists.

Gap junctions. (A) Electron micrograph of a gap junction (arrows) between two liver cells. (B) Gap junctions consist of assemblies of six connexins, which form open channels through the plasma membranes of adjacent cells. (A, Don Fawcett and R. Wood/Photo (more...)
Adhesion between plant cells is mediated by their cell walls rather than by transmembrane proteins. In particular, a specialized pectin-rich region of the cell wall called the middle lamella acts as a glue to hold adjacent cells together. Because of the rigidity of plant cell walls, stable associations between plant cells do not require the formation of cytoskeletal links, such as those provided by the desmosomes and adherens junctions of animal cells. However, adjacent plant cells communicate with each other through cytoplasmic connections called plasmodesmata (singular, plasmodesma), which function analogously to animal cell gap junctions.
Despite their similarities in function, plasmodesmata are structurally unrelated to gap junctions. At each plasmodesma the plasma membrane of one cell is continuous with that of its neighbor, creating an open channel between the two cytosols (Figure 12.67). An extension of the smooth endoplasmic reticulum passes through the pore, leaving a ring of surrounding cytoplasm through which ions and small molecules are able to pass freely between the cells. In addition, plasmodesmata can expand in response to appropriate stimuli, permitting the regulated passage of macromolecules between adjacent cells. Plasmodesmata may thus play a key role in plant development by controlling the trafficking of regulatory molecules, such as transcription factors or RNAs, between cells.

Plasmodesmata. (A) Electron micrograph of plasmodesmata (arrows). (B) At plasmodesmata, the plasma membranes of neighboring cells are continuous, forming cytoplasmic channels through the adjacent cell walls. An extension of the endoplasmic reticulum usually (more...)
By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Your browsing activity is empty.
Activity recording is turned off.
See more...