

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Institute of Medicine (US) Forum on Microbial Threats. The Causes and Impacts of Neglected Tropical and Zoonotic Diseases: Opportunities for Integrated Intervention Strategies. Washington (DC): National Academies Press (US); 2011.

The Causes and Impacts of Neglected Tropical and Zoonotic Diseases: Opportunities for Integrated Intervention Strategies.
Show detailsIn this study, we hypothesize that the worldwide distribution of cognitive ability is determined in part by variation in the intensity of infectious diseases. From an energetics standpoint, a developing human will have difficulty building a brain and fighting off infectious diseases at the same time, as both are very metabolically costly tasks. Using three measures of average national intelligence quotient (IQ), we found that the zero-order correlation between average IQ and parasite stress ranges from r = -0.76 to r = -0.82 (p < 0.0001). These correlations are robust worldwide, as well as within five of six world regions. Infectious disease remains the most powerful predictor of average national IQ when temperature, distance from Africa, gross domestic product per capita and several measures of education are controlled for. These findings suggest that the Flynn effect may be caused in part by the decrease in the intensity of infectious diseases as nations develop.
1. Introduction
Since the first publication of quantitative data on average national intelligence quotient (IQ) scores (Lynn & Vanhanen, 2001, 2002, 2006), five empirical studies have attempted to explain the global distribution of variation in intelligence. Barber (2005) hypothesized that IQ—like many other psychological traits—is a highly plastic trait that may increase onto genetically as the rewards of higher intelligence increase, and with exposure to education and other cognitively demanding environments such as non-agricultural labour. He reported that, across 81 nations, average national IQ correlated with enrolment in secondary school (r = 0.72), illiteracy (r = -0.71), agricultural labour (r = -0.70) and gross national product (r = 0.54). He also proposed that health and nutrition may affect intelligence, and found that average national IQ correlated negatively with rates of low birth weight (r = -0.48) and with infant mortality (r = -0.34). While it is plausible that formal education increases intelligence, Barber (2005) admits that it is not possible to determine from the data he used whether the correlation between education and intelligence is owing to education increasing intelligence or whether more intelligent individuals seek more education. Research has shown this relationship to be intractable (reviewed in Ceci, 1991). The same direction-of-causation ambiguity is true for agricultural labour (Barber, 2005). We agree with Barber's assertions that health and nutrition may affect intelligence, although the variables he studied—low birth weight and infant mortality—are probably rather incomplete measures of these factors.
Lynn (1991) and Rushton (1995, 2000) proposed that temperature and climate provide important Darwinian selective pressures for intelligence, with cold climates selecting for higher intelligence, because low temperatures provide more fitness-related problems for humans that must be solved through cognitively demanding means, and through more complex social organization. Templer & Arikawa (2006) tested and supported predictions of this proposal in a cross-national study and found that average IQ correlated significantly with winter high temperature (r = -0.76), winter low temperature (r = -0.66), summer high temperature (r = -0.31) and summer low temperature (r = -0.41). Templer & Arikawa (2006) also found that average IQ correlated significantly with average skin darkness (r = -0.92). The authors offered little explanation of why this trend exists, except that they believed skin colour was related to exposure to certain climates over evolutionary time.
Kanazawa (2004) hypothesized that intelligence evolved as a domain-specific psychological adaptation to deal with environments that are evolutionarily novel. This hypothesis was tested and supported at the crossnational level (Kanazawa 2008). Results showed that distance from three points in or near central Africa—the evolutionary origin of humans—correlated positively with average national IQ (0° E, 0° N, r = 0.45; South Africa, r = 0.53; Ethiopia, r = 0.22). Kanazawa (2008) did not offer his findings as an alternative to those of Templer & Arikawa (2006), but, rather, as complementary to them. Wicherts et al. (2010a) and Borsboom & Dolan (2006) heavily criticized Kanazawa's hypothesis; for reasons they give in detail, we seriously question the ability of linear distance from sub-Saharan Africa to measure evolutionary novelty, undermining the foundation of Kanazawa's hypothesis.
Saadat (2008) and Woodley (2009) suggested that inbreeding depression and associated reduced phenotypic quality is a cause of the variation in cognitive ability across the world. They found cross-national correlations of r = -0.77 (n = 35, p < 0.0001) and r = -0.62 (n = 71, p < 0.01), respectively, between average IQ and measures of inbreeding. Woodley (2009), however, noted that rates of consanguineous marriage itself may not account for the magnitude of this variation because (i) the statistical significance of the effect disappears when education and gross domestic product (GDP) are controlled for, and (ii) the effect of inbreeding on intelligence had previously been shown to be relatively small.
Here, we offer a new hypothesis—the parasite-stress hypothesis—to explain the worldwide distribution of intelligence. The brain is the most complex and costly organ in the human body. In human newborns, the brain demands 87 per cent of the body's metabolic budget, 44 per cent at age five, 34 per cent at age ten, and 23 per cent and 27 per cent for adult males and females, respectively (Holliday, 1986). Presumably, if an individual cannot meet these energetic demands while the brain is growing and developing, the brain's growth and developmental stability will suffer. Lynn (1990, 1993) has argued that nutrition is vital to high degrees of mental development. Lynn (1990) suggested that nutrition may account for the Flynn effect (large increases in IQ over short periods of time as nations develop; Flynn, 1987), and later (Lynn, 1993) reviewed evidence showing that undernourished children have smaller heads, smaller brains and lower psychometric intelligence than sufficiently nourished children.
Parasitic infection affects the body, and hence the brain, energetically in four ways. (i) Some parasitic organisms feed on the host's tissues: the loss must be replaced at energetic cost to the host. Such organisms notably include flukes and many kinds of bacteria. (ii) Some parasites inhabit the intestinal tract or cause diarrhoea, limiting the host's intake of otherwise available nutrients. These notably include tapeworms, bacteria, giardia and amoebae. (iii) Viruses use the host's cellular machinery and macromolecules to reproduce themselves, at the energetic expense of the host. (iv) The host must activate its immune system to fight off the infection, at energetic expense. Of these, diarrhoeal diseases may impose the most serious cost on their hosts' energy budget. First, diarrhoeal diseases are the most common category of disease on every continent, and are one of the two top killers of children under five, accounting for 16 to 17 per cent of all of these deaths worldwide (WHO, 2004a). Second, diarrhoea can prevent the body from accessing any nutrients at all. If exposed to diarrhoeal diseases during their first five years, individuals may experience lifelong detrimental effects to their brain development, and thus intelligence. Parasites may negatively affect cognitive function in other ways, such as by infecting the brain directly, but we focus only on energetic costs.
The worldwide distribution of parasites is well known. Disease-causing organisms of humans are more prevalent in equatorial regions of the world and become less prevalent as latitude increases. Ecological factors contributing to this distribution include mean annual temperature, monthly temperature range and precipitation (e.g. Guernier et al., 2004). Similar trends of parasite distribution have been shown in other host species (e.g. Møller, 1998).
Many studies have shown a negative relationship between intestinal helminth infection and cognitive ability (reviewed in Watkins & Pollitt, 1997; see also Dickson et al., 2000). Although several hypotheses have been proposed to explain this phenomenon, none have considered intestinal worms in the larger context of all parasitic infection, nor have they considered fully the energetic cost of infection and its consequences on the brain. Other studies have shown relationships between helminth infection and economic and educational factors that are related to intelligence. For example, Bleakley (2007) studied the effects of eradication of hookworm in the southern US during the early twentieth century, and found that areas where hookworm infections had been greatly reduced had higher average incomes after treatment than areas that had not received treatment. Jardin-Botelho et al. (2008) found that Brazilian children infected with hookworm performed more poorly on cognitive tests than uninfected children, and that children infected with more than one type of intestinal helminth performed more poorly than children infected with only one.
Thus, from the parasite-stress hypothesis, we predict that average national intelligence will correlate significantly and negatively with rates of infectious disease, and that infectious disease will remain an important predictor of average national intelligence when other variables are controlled for. It is the purpose of this study to introduce this hypothesis to describe the worldwide variation in intelligence, and to provide some supportive evidence using correlations and linear modeling techniques.
2. Material and Methods
National average intelligence was taken from Lynn & Vanhanen (2006), who present their methods in detail. IQ was measured directly in 113 nations, and estimated for 79 more nations by averaging the IQs of nearby nations with known IQ. These estimates were validated by Lynn & Vanhanen (2006) by comparing them to actual measurements of IQ in the same nations. At least two studies have presented evidence of validation for these data (Lynn & Mikk, 2007; Rindermann, 2007), by showing strong positive correlations between Lynn & Vanhanen's (2006) national IQ scores and other measures of cognitive ability. Wicherts et al. (2010b) have criticized Lynn & Vanhanen's (2006) estimates of IQ in sub-Saharan African nations on the grounds that the selection criteria used to include studies in these estimations did not produce national IQ scores that were representative. Using selection criteria that they argue are more appropriate, Wicherts et al. (2010b) proposed new average IQ values for 17 nations in sub-Saharan Africa (but see also Lynn & Meisenberg, 2010). Analyses will be performed using three datasets: Lynn & Vanhanen's (2006) original data, including estimates (LVE; mean = 84, median = 84.2, s.d. = 11.8); Lynn & Vanhanen's (2006) data using collected data only (LVCD; mean = 86.7, median = 87.5, s.d. = 11.9); and Wicherts et al.'s (2010b) revisions of Lynn & Vanhanen's (2006) data with estimates (WEAM; mean = 85.0, median = 85, s.d. = 11.0).
As a measure of infectious disease levels for each nation, disability-adjusted life years lost (DALY) owing to infectious disease were used (WHO, 2004b). This measure combines years of life lost and years spent disabled owing to 28 representative and important human diseases, including tetanus, malaria, tuberculosis, hepatitis, syphilis and leishmaniasis, such that one DALY equals one healthy year of life lost per 100 000 people. Although other cross-national measures of disease exist, we believe this to be the best for our study because (i) data exist for most countries of the world (n = 192), and (ii) this variable is a reasonable measure of the physiological costs of infectious disease, which concerns the parasite-stress hypothesis applied to cognitive ability. The DALY infectious-disease measure correlates strongly with other measures of human infectious disease (e.g. Thornhill et al., 2009). This variable was log-transformed owing to an extreme skew to the right (mean = 3.36, median = 3.28, s.d. = 0.761).
As an independent measure of nutrient stress, DALY owing to nutritional deficiencies (WHO, 2004b) were used. This calculation includes mortality and healthy years lost owing to protein-energy malnutrition, iodine deficiency, vitamin A deficiency and iron-deficiency anaemia. This variable was log-transformed owing to an extreme skew to the right (mean = 2.59, median = 2.65, s.d. = 0.49).
Average winter high temperatures (mean = 15.6, median = 17, s.d. = 12.5) were taken from Templer & Arikawa (2006). Although they used four intercorrelated temperature variables in their analysis, they reported that average winter high temperature was the best predictor of IQ of the four, so we used it.
Although Templer & Arikawa (2006) found a positive relationship between IQ and skin darkness, we will not use skin darkness in our analyses for three reasons: (i) although evidence suggests that skin darkness is a measure of historical infectious disease intensity over evolutionary time, it is unclear exactly what kind of infectious diseases it is indicative of (see discussion); (ii) Templer & Arikawa (2006) argued that the relationship between skin darkness and IQ is not causal; and (iii) Templer & Arikawa (2006) did not sufficiently explain why the association between intelligence and skin darkness exists. Without a reasonable theoretical framework for this association, we did not feel it was appropriate to compare it with other variables for which there is a better theoretical rationale.
Literacy rates (mean = 87.6, median = 96, s.d. = 15.9) were taken from World Bank (2008). This variable is defined as the percentage of the population aged 15 years and older who have at least a basic proficiency at reading. Data were from the most recent year available for each nation between 1990 and 2007. Barber (2005) used data from 1976 for his analysis of literacy because that was the average year in which IQ data were collected for all countries. We felt that a more recent date was more appropriate, however, because the IQ scores for each country we used had been modified based on recorded trends of the Flynn effect to reflect the expected modern IQ score (Lynn & Vanhanen, 2006).
Enrolment in secondary school (mean = 29.0, median = 28.5, s.d. = 16.8), completion of secondary school (mean = 12.0, median = 10.5, s.d. = 9.25) and average years of education (AVED; mean = 6.17, median = 5.81, s.d. = 2.89) were taken from Barro & Lee (2001). These numbers represent the percentage of the population aged 25 and older who have attended some or all of secondary school, and the average number of years of schooling in the population. Data were used for the most recent years available after 1990. Data that were only available for years prior to 1990 were omitted.
Data of GDP per capita in US dollars was taken from the World factbook (CIA, 2007), and were log-transformed for normality (mean = 3.81, median = 3.85, s.d. = 0.53).
Distance from central Africa, or the human environment of evolutionary adaptedness (EEA), was calculated using the Pythagorean Theorem, as done by Kanazawa (2008). Kanazawa used three points at the corners of sub-Saharan Africa, so theoretically any point within this triangle should be a valid centre point from which to calculate distance. We selected 258 latitude, 2258 longitude, which is in the approximate centre of this area, and calculated distance from this point using the same methods as Kanazawa. Despite our own criticism of this variable and that of others (Wicherts et al., 2010a), we included this variable in the interest of thoroughness. This variable was log-transformed for normality (mean = 4.0, median = 3.97, s.d. = 0.710).
Percentage consanguineous marriages was not used in this study because it was the conclusion of Woodley (2009) that this variable is unlikely to account for the worldwide variation in intelligence. Additionally, consanguineous marriage and associated inbreeding may be a strategy for maintaining coadapted gene complexes that defend against local infectious diseases (Denic & Nicholls, 2007; Fincher & Thornhill, 2008).
All analyses were performed using JMP 8.0.2 statistical software.
3. Results
Log DALY owing to infectious disease and average national IQ correlated at r = -0.82 (n = 107, p < 0.0001) using LVCD, r = -0.82 (n = 184, p < 0.0001; figure A3-1) using LVE and r = -0.76 (n = 184, p < 0.0001; figure A3-1) using WEAM. Zero-order correlations were also performed for each of Murdock's (1949) six world regions (see table A3-2). A hierarchical linear model (HLM) was also performed to determine whether this relationship is consistent across the six regions, finding that it is (LVE: R2 = 0.78, p < 0.0001, n = 184; LVCD: R2 = 0.77, p < 0.0001, n = 107; WEAM: R2= 0.68, p < 0.0001, n = 184).
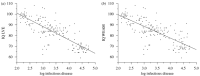
FIGURE A3-1
Log DALY owing to infectious disease and average national IQ correlate (a) at r = -0.82 (LVE) and (b) at r = -0.76 (WEAM; n = 184, p < 0.0001). The line is the least-squares line through the points.
TABLE A3-2
Zero-order correlations between average national intelligence and log DALY owing to infectious disease within each of Murdock's (1949) six world regions. Values not in parentheses used LVE, values in parentheses used LVCD and values in square brackets (more...)
Log DALY owing to nutritional deficiencies and IQ correlated at r = -0.72 (n = 184, p < 0.0001). Log DALY owing to infectious disease and log DALY owing to nutritional deficiencies correlated at r = 0.89 (n = 192, p < 0.0001). The partial correlation between IQ and DALY owing to nutritional deficiencies with the effects of DALY owing to infectious disease removed was near zero (r = 0.028; n = 184, p = 0.71), while the partial correlation between IQ and DALY owing to infectious disease with the effects of DALY owing to nutritional deficiencies removed remained strong (r = -0.56; n = 184, p < 0.0001). See table A3-1 for other zero-order correlations. The correlations between these variables and IQ are very similar across the three measures of IQ (see the electronic supplementary material, tables A3-S1 and A3-S2).
TABLE A3-1
Zero-order correlations among average national IQ (LVE), log DALY owing to infectious disease, average winter high temperature, distance from EEA, literacy, average years of education (AVED), % enrolling in secondary education, % completing all secondary (more...)
TABLE A3-S-1
Zero-order correlations among average National IQ (LVCD), log DALY infectious disease, average winter high temperature, distance from EEA, literacy, average years of education (AVED), % enrolling in secondary education, % completing all secondary education, (more...)
TABLE A3-S-2
Zero-order correlations among average National (WEAM), log DALY infectious disease, average winter high temperature, distance from EEA, literacy, average years of education (AVED), % enrolling in secondary education, % completing all secondary education, (more...)
To select which, if any, education and wealth variables to include in a multiple regression analysis, partial correlations were performed independently between literacy, enrolment in secondary school, completion of secondary school and AVED, and average national IQ, with the effects of infectious disease removed. If a variable was no longer significant when the effects of infectious disease were removed, it was not included in the multiple regression. Only WEAM and LVE IQ measures were used for these multivariate analyses in order to have a sample size large enough to make inferences about the individual contributions of each variable. When the effects of log DALY owing to infectious disease were removed, the correlation between IQ and literacy was r = 0.15 (n = 113, p = 0.094) using LVE and r = 0.16 (n = 113, p = 0.1087) using WEAM; IQ and some secondary education was r = 0.093 (n = 120, p = 0.32) using LVE, and r = 0.23 (n = 120, p = 0.049) using WEAM; IQ and completion of secondary education was r = 0.17 (n = 110, p = 0.08) using LVE and r = 0.23 (n = 110, p = 0.030) using WEAM; IQ and AVED was r = 0.23 (n = 127, p = 0.0084) using LVE and r = 0.30 (n = 127, p = 0.0005) using WEAM; and IQ and GDP was r = 0.054 (n = 184, p = 0.46) using LVE and r = 0.036 (n = 184, p = 0.61) using WEAM. AVED was the best predictor of IQ when the effects of infectious disease were removed for both measures of IQ, so this education variable was used in regressions. As such, AVED will have the best chance of all the education variables at being significant in the multiple regression. Although GDP was not a statistically significant predictor of IQ when the effects of infectious disease were removed, and the partial correlation coefficients were well below 0.1, we included this variable in some models at the request of a reviewer (table A3-3).
TABLE A3-3
Multiple regression analyses predicting average national intelligence using LVE and WEAM (in parentheses where different) by log DALY owing to infectious disease, log distance from EEA, average winter high temperature, average years of education (AVED) (more...)
In a multiple linear regression, average national IQ (LVE and WEAM) was predicted using infectious disease, average winter high temperature, distance from sub-Saharan Africa, AVED and GDP (see table A3-3 for model details). Significant predictors in this model were infectious disease, distance from Africa and winter high temperature. AVED was not significant. When GDP was removed from this model, virtually identical patterns emerged (see the electronic supplementary material, table A3-S3 for model details).
TABLE A3-S-3
Multiple regression analyses predicting average national intelligence using LVE and WEAM (in parentheses when different) by log DALY infectious disease, log distance from EEA, average winter high temperature, and average years of education (AVED).
4. Discussion
The negative relationship between infectious disease and IQ was statistically significant at the national level both worldwide and within five of Murdock's (1949) six world regions. All analyses showed that infectious disease was a significant predictor of average national IQ, whether using either of Lynn & Vanhanen's (2006) two datasets or Wicherts et al.'s (2010b) data. The zero-order correlation between DALY owing to infectious disease and average national IQ was higher than that of any other variable for which there is a previously proposed causal explanation. The world regions analysis showed that the international pattern is repeated within five of the six regions despite a region's generally similar cultural history. The only world region in which this relationship was not significant was South America. This exception may be owing to the presence of several outliers. The group of conspicuous outliers in which IQ was much lower than expected in the worldwide trend (figure A3-1) are all Caribbean countries (St Lucia, Dominica, St Kitts and Nevis, Antigua and Barbuda, Grenada, St Vincent and Grenadines, and Jamaica), which represent 4 of 23 nations in the South America analysis (St Lucia, Dominica, Grenada, and St Vincent and the Grenadines). Because these outliers are in the same geographical location, it is possible that local parasites that are not included in the DALY owing to infectious disease variable are causing these outliers. HLM analysis shows that, despite the nonsignificance of the correlation between IQ and infectious disease within South American nations, this trend is significant overall across Murdock's (1949) six world areas.
Nutritional stress correlated with average national IQ (r = -0.72), but this relationship was not significant when the effects of infectious disease were removed. This supports the suggested link between intelligence and nutrition. Given the energetic cost of infectious disease, individuals who are burdened with parasites may be more likely to be affected by nutritional deficiencies. Likewise, individuals who are suffering from nutritional deficiencies may be less able to mount an effective immune response.
Multiple regression shows that, of infectious disease, temperature, evolutionary novelty and AVED, infectious disease is the best predictor of intelligence by a large margin. The effects of years of education are not significant, while temperature and evolutionary novelty seem to have distinct predictive power beyond infectious disease. Although this model cannot rule out the independent effect of distance from central Africa, this effect is difficult to interpret because of the doubt cast on the theory underlying this variable (Wicherts et al., 2010a). Although the effects of education and GDP per capita are not statistically significant when other factors are controlled for, this is not to say that these factors are not involved. A nation of more intelligent individuals is likely to produce a higher GDP, but a wealthier nation is also more able to pay for public education, as well as public medical and sanitation services. An indirect link between education and intelligence may also exist, as a better-educated population may be more interested in public health measures—leading to increased IQ by reducing information about germ theory and hygiene. These sources of endogeneity must be considered when interpreting our findings (and see below). It should also be mentioned that we are not arguing that global variation in intelligence is only caused by parasite stress. Rather, variation in intelligence is probably caused by a variety of factors, including those we have mentioned here as well as factors that are yet unknown.
If the general pathway we propose is correct, there are two plausible mechanisms by which a trade-off in allocation of energy to immune function versus brain development and maintenance may occur. First, parasitic infection may intermittently cause the redirection of energy away from brain development. In this case, during periods of infection, the brain receives fewer energetic resources, but this allocation to brain function will return to pre-infection levels during healthy periods.
During periods of infection, whatever aspects of the brain that are growing and developing will suffer reduced phenotypic quality. Second, exposure to infectious agents may cause a developmental pathway that permanently invests more energy into immune function at the expense of brain growth. In this scenario, large amounts of energy would be allocated into immune function during periods of health, as opposed to only redirecting energy during periods of infection. This could operate through a variety of mechanisms. A plausible mechanism is that higher investment in immune system is triggered by individual exposure to infectious disease at some point during ontogeny. This may include triggering from exposure to maternal antibodies while in utero.
We also propose a complementary hypothesis that may explain some of the effects of infectious disease on intelligence. As we mentioned, it is possible that a conditional developmental pathway exists that invests more energy into the immune system at the expense of brain development. In an environment where there has consistently been a high metabolic cost associated with parasitic infection, selection would not favour the maintenance of a phenotypically plastic trait. That is, the conditional strategy of allocating more energy into brain development during periods of health would be lost, evolutionarily, if periods of health were rare. Peoples living in areas of consistently high prevalence of infectious disease over evolutionary time thus may possess adaptations that favour high obligatory investment in immune function at the expense of other metabolically expensive traits such as intelligence. Data do not currently exist on temporal variation of the severity of infectious disease across the world over human history. For genetically distinct adaptations in intelligence to exist based on this principle, parasite levels must be quite consistent over evolutionary time. If this is not the case, then selection would maintain investment in the immune system and in the brain as a plastic (as opposed to static) trait. The Flynn effect (Flynn, 1987) indicates that conditional developmental causes must be at work at least in part. Large increases in intelligence across a few generations cannot be attributed to genetic differences caused by evolutionary processes. Hence, it does not seem probable that region-specific genetic adaptations are the primary cause of the worldwide variation in intelligence.
Our findings suggest that the heritable variation in intelligence may come from two sources: brain structure and immune system quality. Thus, two individuals may possess identical genes for brain structure, but have different IQ owing to differences in immune system quality reflecting their personal allocation of energy into brain development versus immunity.
Our findings are consistent with a number of other findings in the literature. In particular, the Flynn effect (Flynn 1987) demands that any hypothesis regarding the worldwide variation and distribution of intelligence must be able to account for some factor that allows for large IQ gains over time spans seemingly too short to be attributed to evolution by natural selection. The parasite-stress hypothesis allows for such a factor in the form of reduced parasitic infection. As societies become modernized, decreased parasite stress may occur through multiple pathways. As national wealth increases, medicine, vaccinations and potable water can be purchased by both the government and by individuals. Moreover, there is cross-national evidence that, as democratization increases, there are corresponding increases in public health legislation and infrastructure. Democratization also increases levels of education, better allowing individuals to seek out and understand information that reduces parasitic infection (Thornhill et al., 2009). This source of endogeneity is not a flaw, but a prediction of our hypothesis.
Mackintosh (2001) presented comprehensive evidence that skin darkness and the associated cellular components (e.g. melanocytes) have an important role in defending against infectious disease. Moreover, Manning et al. (2003) found that, in sub-Saharan Africa, rates of HIV infection were negatively associated with skin darkness. Manning et al. (2003) attributed this relationship in part to lower infection rates of other parasites, especially bacteria and fungi, that lead to tissue damage in the genital tract and hence increased opportunity for contracting HIV. Templer & Arikawa (2006) concluded that, despite the strong negative correlation between skin colour and average national IQ, there must be an unknown mediating factor accounting for both because there is no obvious reason for skin darkness to reduce IQ. Given the previous research linking skin colour to infectious disease (Mackintosh, 2001; Manning et al., 2003), the unknown factor linking skin colour and IQ may be infectious disease.
Several studies have shown a positive relationship between IQ and body symmetry (e.g. Furlow et al., 1997; Prokosch et al., 2005; Bates, 2007; Penke et al., 2009; but see also Johnson et al., 2008). There is evidence that body symmetry is a measure of developmental stability, an important component of which is owing to reduced contact with infectious disease (Thornhill & Møller, 1997). Our study suggests that IQ and body symmetry correlate because they are both affected negatively by exposure to high infectious disease. Individuals who are exposed to infectious disease may have many aspects of their body develop imperfectly, including the brain, negatively affecting both their body symmetry and cognitive ability. Indeed, recent research indicates that there is a positive relationship between body asymmetry and atypical brain asymmetries (Yeo et al., 2007).
The hygiene hypothesis proposes that some autoimmune diseases may be caused by low exposure to pathogens during ontogeny (e.g. Strachan, 1989). Previous studies of individual differences have shown that intelligence correlated positively with the frequency of asthma and allergies (reviewed in Jensen & Sinha, 1993). According to the parasite-stress hypothesis, high intelligence is allowed in part by low exposure to infectious disease. Thus the relationship between intelligence and autoimmune diseases, such as asthma and allergies (reviewed in Gangal & Chowgule, 2009), is probably mediated through exposure to infectious disease. We predict that this positive relationship between IQ and autoimmune diseases will also be robust across nations, and that it will be mediated by infectious disease.
Although our results support our predictions, further studies must be done to establish causation. Longitudinal methods could be used to test this hypothesis on the individual level. Children's IQ could be measured at an early age and remeasured later in life, while monitoring for infectious diseases throughout childhood. This would not only provide another test of our hypothesis, but may be able to determine the effects of individual infectious diseases on cognitive development. Additionally, it could be determined which, if either, trade-off mechanism we discussed is responsible for the detrimental effects of infectious disease on intelligence. Both may operate but with geographical differences based on the consistency of infectious disease over time. As nations develop, they could be monitored for declining rates of parasitic infection to determine (i) whether this corresponds with elevated IQ and (ii) whether any IQ gain is sufficient to account for the Flynn effect.
The authors thank Rachael Falcon, Steve Gangestad, Angela Hung, Kenneth Letendre, Paul Watson and Rhiannon West for their comments and suggestions. Three anonymous reviewers provided helpful and polite criticism.
References
- Barber N. Educational and ecological correlates of IQ: a cross-national investigation. Intelligence. 2005;33:273–284. [CrossRef]
- Barro RJ, Lee J. International data on educational attainment: updates and implications. Oxford Econ. Pap. 2001;53:541–563. [CrossRef]
- Bates TC. Fluctuating asymmetry and intelligence. Intelligence. 2007;35:41–46. [CrossRef]
- Bleakley H. Disease and development: evidence from hookworm eradication in the American south. Q. J. Econ. 2007;122:73–117. [PMC free article: PMC3800113] [PubMed: 24146438]
- Borsboom D, Dolan CV. Why g is not an adaptation: a comment on Kanazawa (2004). Psychol. Rev. 2006;113:433–437. [PubMed: 16637768] [CrossRef]
- Ceci SJ. How much does schooling influence general intelligence and its cognitive components? A reassessment of the evidence. Dev. Psychol. 1991;27:703–722. [CrossRef]
- CIA. World factbook. 2007. See https://www
.cia.gov/library /publications/the-world-factbook. - Denic S, Nicholls MG. Genetic benefits of consanguinity through selection of genotypes protective against malaria. Hum. Biol. 2007;79:145–158. [PubMed: 18027811] [CrossRef]
- Dickson R, Awasthi S, Williamson P, Demellweek C, Garner P. Effects of treatment for intestinal helminth infection on growth and cognitive performance in children: systematic review of randomized trials. Br. Med. J. 2000;320:1697–1701. [PMC free article: PMC27412] [PubMed: 10864543] [CrossRef]
- Fincher CL, Thornhill R. A parasite-driven wedge: infectious diseases may explain language and other biodiversity. Oikos. 2008;117:1289–1297. [CrossRef]
- Flynn JR. Massive IQ gains in 14 nations: what IQ tests really measure. Psychol. Bull. 1987;101:171–191. [CrossRef]
- Furlow FB, Armijo-Prewitt T, Gangestad SW, Thornhill R. Fluctuating asymmetry and psychometric intelligence. Proc. R. Soc. Lond. B. 1997;264:823–829. [PMC free article: PMC1688437] [PubMed: 9265189] [CrossRef]
- Gangal SV, Chowgule R. Infections in early life and susceptibility to allergic diseases: relevance of hygiene hypothesis. Curr. Sci. India. 2009;96:784–793.
- Guernier V, Hochberg ME, Guégan J. Ecology drives the worldwide distribution of human diseases. PLOS Biol. 2004;2:740–746. [PMC free article: PMC423130] [PubMed: 15208708]
- Holliday MA. Body composition and energy needs during growth, Human growth: a comprehensive treatise. Falkner F, Tanner JM, editors. Vol. 2. New York, NY: Plenum; 1986. pp. 101–117.
- Jardin-Botelho A, Raff S, Rodrigues RA, Hoffman HJ, Diemert JH, Correa-Oliviera R, Bethony JM, Gazzinelli MF. Hookworm, Ascaris lumbricoides infection and polyparasitism associated with poor cognitive performance in Brazilian schoolchildren. Trop. Med. Int. Health. 2008;13:994–1004. [PubMed: 18627581]
- Jensen AR, Sinha SN. Physical correlates of human intelligence, Biological approaches to the study of human intelligence. Vernon PA, editor. Norwood, NJ: Ablex; 1993. pp. 139–242.
- Johnson W, Segal NL, Bouchard TJ Jr. Fluctuating asymmetry and general intelligence: no genetic or phenotypic association. Intelligence. 2008;36:279–288. [CrossRef]
- Kanazawa S. General intelligence as a domain-specific adaptation. Psychol. Rev. 2004;111:512–523. [PubMed: 15065920] [CrossRef]
- Kanazawa S. Temperature and evolutionary novelty as forces behind the evolution of general intelligence. Intelligence. 2008;36:99–108. [CrossRef]
- Lynn R. The role of nutrition in secular increases in intelligence. Person. Indiv. Differ. 1990;11:263–285.
- Lynn R. The evolution of racial differences in intelligence. Mankind Quart. 1991;32:99–121.
- Lynn R. Nutrition and intelligence, Biological approaches to the study of human intelligence. Vernon PA, editor. Norwood, NJ: Ablex; 1993. pp. 243–258.
- Lynn R, Meisenberg G. The average IQ of sub-Saharan Africans: comments of Wicherts, Dolan, and van der Maas. Intelligence. 2010;38:21–29. [CrossRef]
- Lynn R, Mikk J. National differences in intelligence and educational attainment. Intelligence. 2007;35:115–121. [CrossRef]
- Lynn R, Vanhanen T. National IQ and economic development: a study of eighty-one nations. Mankind Quart. 2001;41:415–435.
- Lynn R, Vanhanen T. IQ and the wealth of nations. Westport, CT: Praeger; 2002.
- Lynn R, Vanhanen T. IQ and global inequality. Augusta, GA: Washington Summit; 2006.
- Mackintosh JA. The antimicrobial properties of malanocytes, melanosomes and melanin and the evolution of black skin. J. Theor. Biol. 2001;211:101–113. [PubMed: 11419954] [CrossRef]
- Manning JT, Bundred PE, Henzi P. Melanin and HIV in sub-Saharan Africa. J. Theor. Biol. 2003;223:131–133. [PubMed: 12782121] [CrossRef]
- Møller AP. Evidence of larger impact of parasites on hosts in the tropics: investment in immune function within and outside the tropics. Oikos. 1998;82:265–270. [CrossRef]
- Murdock GP. Social structure. New York, NY: Macmillan; 1949.
- Penke L, Bates TC, Gow AJ, Pattie A, Starr JM, Jones BC, Perrett DI, Deary IJ. Symmetric faces are a sign of successful cognitive aging. Evol. Hum. Behav. 2009;30:429–437. [CrossRef]
- Prokosch MD, Yeo RA, Miller GF. Intelligence tests with higher g loadings show higher correlations with body symmetry: evidence for a general fitness factor mediated by developmental stability. Intelligence. 2005;33:203–213. [CrossRef]
- Rindermann H. The g-factor of international cognitive ability comparisons: the homogeneity of results in PISA, TIMSS, PIRLS and IQ-tests across nations. Eur. J. Personality. 2007;21:667–706. [CrossRef]
- Rushton JP. Race, evolution, and behavior: a life history perspective. New Brunswick, NJ: Transaction; 1995.
- Rushton JP. Race, evolution, and behavior: a life history perspective. 2nd special abridged. Port Huron, MI: Charles Darwin Research Institute; 2000.
- Saadat M. Consanguinity and national IQ scores. J. Epidemiol. Commun. H. 2008;62:566–567. [PubMed: 18477759] [CrossRef]
- Strachan DP. Hay-fever, hygiene, and household size. Brit. Med. J. 1989;299:1259–1260. [PMC free article: PMC1838109] [PubMed: 2513902] [CrossRef]
- Templer DI, Arikawa H. Temperature, skin color, per capita income, and IQ: an international perspective. Intelligence. 2006;34:121–139. [CrossRef]
- Thornhill R, Møller AP. Developmental stability, disease and medicine. Biol. Rev. 1997;72:497–548. [PubMed: 9375532] [CrossRef]
- Thornhill R, Fincher CL, Aran D. Parasites, democratization, and the liberalization of values across contemporary countries. Biol. Rev. 2009;84:113–131. [PubMed: 19046399] [CrossRef]
- Watkins WE, Pollitt E. ‘Stupidity or worms’: do intestinal worms impair mental performance. Psychol. Bull. 1997;121:171–191. [PubMed: 9100486] [CrossRef]
- Wicherts JM, Borsboom D, Dolan CV. Why national IQs do not support evolutionary theories of intelligence. Person. Indiv. Differ. 2010;48:91–96. [CrossRef]
- Wicherts JM, Dolan CV, van der Maas HLJ. A systematic literature review of the average IQ of sub-Saharan Africans. Intelligence. 2010;38:1–20. [CrossRef]
- Woodley MA. Inbreeding depression and IQ in a study of 72 countries. Intelligence. 2009;37:268–276. [CrossRef]
- World Bank. The world at a glance: key development indicators from the World Bank. 2008. See http://data
.worldbank.org. - WHO. Global burden of disease: 2004 update. Geneva, Switzerland: World Health Organization; 2004.
- WHO. World health report 2004. Geneva, Switzerland: World Health Organization; 2004.
- Yeo RA, Gangestad SW, Thoma RJ. Developmental instability and individual variation in brain development: implications for the origin of neurodevelopmental disorders. Curr. Dir. Psychol. Sci. 2007;16:245–249. [CrossRef]
Footnotes
- 2
Reprinted from Eppig, C., Fincher, C. L., and Thornhill R. (2010). Parasite prevalence and the worldwide distribution of cognitive ability, Proceedings of the Royal Society Biological Sciences, with permission from The Royal Society.
- 3
Author for correspondence ( ude.mnu@gippec).
- PARASITE PREVALENCE AND THE WORLDWIDE DISTRIBUTION OF COGNITIVE ABILITY - The Ca...PARASITE PREVALENCE AND THE WORLDWIDE DISTRIBUTION OF COGNITIVE ABILITY - The Causes and Impacts of Neglected Tropical and Zoonotic Diseases
Your browsing activity is empty.
Activity recording is turned off.
See more...