

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Bermúdez-Rattoni F, editor. Neural Plasticity and Memory: From Genes to Brain Imaging. Boca Raton (FL): CRC Press/Taylor & Francis; 2007.

Abstract
The advent of human brain imaging techniques (PET, fMRI) has allowed previously unimaginable examination of human brain functions including studies of the mechanisms underlying memory for emotional events. Many of these studies have been guided by findings from animal research that identified the amygdala as a key candidate brain region crucial to emotional memory. Evidence from human brain imaging studies has robustly confirmed this conclusion. At the same time, it has expanded our understanding of emotional memory in unsuspected directions. For example, it is now evident that males and females on average do not process memories of the same emotional events in identical ways. This fact is most clear at present regarding the amygdala, for which a sex by hemisphere interaction in its relation to memory has been documented. Whether and how these sex differences influence what is retained in memories of men and women from emotional events is now an important area of research. The evidence to date also makes clear that the long standing and still widespread assumption that subject sex matters little if at all in studies of the neurobiology of emotional memory is no longer tenable and should be abandoned.
15.1. INTRODUCTION
For most of human history, those interested in understanding how brains work possessed few methods to interrogate a healthy human brain without damaging it. This situation changed dramatically about 20 years ago with the advent of imaging techniques such as positron emission tomography (PET) and functional magnetic resonance imaging (fMRI). With these new methods, we are now able to ask questions of the intact, healthy human brain with ever-increasing spatial and temporal resolution. Indeed, so rapid are the advances that those working with these methods struggle to understand how best to utilize this new-found power.
Not surprisingly, scientists interested in the neural mechanisms underlying the influence or emotion on memory (emotional memory) were eager to apply the power of PET and fMRI to their questions. The primary focus of this chapter is on this work. I will also address related work uncovering sex-related influences on other aspects of brain mechanisms of emotional memory storage. A major conclusion from this work is that studies of emotional memory (at least involving human subjects) can no longer safely assume that subject sex will not significantly influence experimental findings and hence conclusions about brain mechanisms.
15.2. THE AMYGDALA: BUILT TO MODULATE
Before discussing this work, it is helpful to appreciate some of the evidence on which the “memory modulation” view of amygdala function rests. Consider first the anatomical connectivity of the primate amygdala. Figure 15.1 comes from a meta-analysis of cortico–cortical connectivity in the monkey by Young and Scannell.1 The analysis conducted by these investigators was agnostic to any particular brain region including the amygdala. Still, they uncovered a striking and unique aspect of the amygdala. Alone among the cortical brain regions examined, the amygdala is possessed of extremely wide connectivity. Furthermore, the vast majority of its connections with the cortex are amygdalofugal — from the amygdala to other brain regions. From this anatomical fact alone, it appears that the amygdala is elegantly and uniquely situated to widely influence cortical function.

FIGURE 15.1
Meta-analysis of cortico–cortico connectivity in the primate brain. Note that the vast majority of amygdala connections are amygdalofugal, suggesting that the amygdala is well positioned to widely influence brain function. (From Young, M.P. and (more...)
This anatomical structure of the amygdala’s connectivity dovetails nicely with a well supported concept of amygdala function, namely that it modulates memory storage processes for emotionally arousing events in other brain regions.2 A very large, diverse, yet cohesive body of evidence indicates that a major amygdala function is to modulate memory storage processes during and after an emotionally arousing event via interactions with endogenous stress hormones released by the events. Across many species, learning tasks, and laboratories, stimulation of the amygdala (and in particular its basolateral complex) potently modulates —enhances or impairs — memory storage processes. Most often, stimulation has been given immediately after learning, allowing the conclusion that the effects of the stimulation on memory resulted from an influence on consolidation processes.2
Extensive and remarkably consistent evidence also indicates that the amygdala’s ability to modulate memory consolidation depends crucially on endogenous stress hormones. Indeed, the same amygdala stimulation may either enhance or impair memory storage, depending on the state of the adrenal glands.2 So strong is the evidence for an interaction between stress hormones and the amygdala in memory that one may fairly conclude that concepts of amygdala function in memory that fail to actively incorporate influences of the body’s hormonal milieu can be considered incomplete at best.
Amygdala function is necessary in essentially all circumstances for peripherally administered drugs and hormones to affect memory. For example, lesions of the amygdala or its key circuitry (e.g., stria terminalis) block the memory-enhancing and -impairing effects of all drugs and hormones tested to date. Even the amnestic effect of general anesthetics is blocked by lesions of the basolateral amygdala.3 Thus the amygdala appears to be a necessary “door” through which all peripherally administered drugs and hormones must go to influence memory storage.
If a major amygdala function is to interact with endogenous stress hormones to influence memory, then we should find a disproportionate effect of amygdala lesions in learning situations that are relatively stress hormone-activating. Cahill and McGaugh4 tested this possibility by examining in rats the effects of amygdala lesions on learning in a series of arousing and presumably hormone-activating tests with their effects on a series of closely matched but relatively non-arousing learning tests. Amygdala lesions impaired memory only in the relatively arousing circumstances. Cahill and McGaugh concluded that “the degree of arousal produced by the unconditioned stimulus, and not the aversive nature per se, determines the level of amygdala involvement (in a learning situation). The amygdala appears to participate in learning especially when the reinforcement is of a highly arousing nature.” The amygdala then is not particularly involved with any particular emotion such as fear, but with arousing learning situations, whether pleasant or unpleasant.
This conclusion is sharply reinforced by four recent human brain imaging studies from four different laboratories, each of which compared the response of the human amygdala to stimuli that varied across the arousing dimension (arousing–calming) and across the valence dimension (pleasant–unpleasant).5–8 All four studies reported that the amygdala responded to the arousing qualities of the stimuli, and not to their valence. As one example, Lewis et al.8 used fMRI to examine the responses of the human amygdala to words that varied according to either their valence or arousal-inducing qualities. Amygdala activity did not correlate with ratings of valence, but correlated significantly with ratings of arousal.
15.3. HUMAN SUBJECT STUDIES RELATING AMYGDALA ACTIVITY TO EMOTIONAL MEMORY
Several studies utilizing human brain imaging examined the amygdala’s role in memory for arousing material. The consistent conclusion from these studies fits very well with the memory modulation view of amygdala function derived from animal research (described above). Amygdala activity while subjects experienced emotional stimuli related significantly to subsequent (usually long-term) memory for the stimuli. However, amygdala activity in the same subjects failed to correlate with subsequent memory for relatively non-arousing stimuli.
In the first of these studies, Cahill et al.9 scanned healthy male subjects with PET for regional cerebral glucose while they viewed either a series of relatively emotionally arousing (negative) films or a matched but much more emotionally neutral set of films. Memory for the films was assessed in an incidental free recall test given 3 weeks later. We found that activity of the right amygdala while viewing a series of emotional films correlated very highly (r = 0.93) with long-term recall of the films but did not relate to recall of the emotionally neutral films.9
This basic finding was subsequently confirmed in three studies from two additional laboratories.10–12 However, unexplained hemispheric asymmetries in the amygdala were evident in each study. We observed that those studies reporting amygdala effects predominantly or exclusively on the right side of the brain involved only male subjects, whereas studies reporting amygdala effects predominantly or exclusively on the left side of the brain involved only female subjects, raising the possibility that subject sex determined, at least in part, the hemispheric lateralization of amygdala function. This conclusion could not, however, be made with confidence on the basis of these studies because they differed along many other dimensions (e.g., type of scanning, type of to-be-remembered material).
15.4. AN ASIDE ON SEX DIFFERENCES IN THE BRAIN
Sex (that is, being male or female) influences brain function to a far greater extent than neuroscience has recognized to date.13 Increasingly we see demonstrations of pronounced neurobiological differences between males and females outside the traditional domain of reproduction in which they are to be expected. Clear sex differences exist in every brain lobe,14 even in “cognitive” brain regions such as the neocortex and hippocampus.
Studies employing human brain imaging techniques report functional sex-related differences in brain correlates of emotional processing,15 facial processing,16,17 working memory,18 auditory processing,19–21 language processing,22,23 and even in visual cortex responsiveness to specific light wavelengths.24 Even cellular mechanisms of neuronal death in cell cultures differ, depending on whether the cells were derived from male or female brains.25 Sex-related differences were also reported in stress hormone responses. For example, Wolf and colleagues recently reported a sex-related difference in the relationship of cortisol to short-term memory.26 They found a negative correlation between the cortisol response to a stressor and memory in their subjects as a whole (men and women considered together), but this effect resulted from a highly significant correlation in men and no such correlation in women.
Finally, sex appears to affect the emotional consequences of amygdala damage in primates. Indications that female monkeys often displayed heightened aggressive behavior following amygdala damage have been in the literature for some time27 — findings in direct opposition to typical effects following amygdala damage in males. Kling28 directly compared the effects of amygdala damage on aggressivity in a group of three female and three male monkeys. He observed that in both his experiment and in the extant literature, “paradoxical” heightened aggressivity after amygdala damage was present only in females. Had potential influences of subject sex been examined starting with the original investigation by Kluver and Bucy,29 we might now have a very different conceptualization of the behavioral consequences of amygdala damage. Within this context, it is clear that studies examining sex-related influences on the amygdala and its relation to memory acquire heightened importance.
15.5. SEX-RELATED INFLUENCES ON AMYGDALA RELATION TO MEMORY FOR EMOTIONAL EVENTS
Given this context, my colleagues and I examined whether subject sex influenced lateralization of the amygdala relationship to long-term memory for emotional material by directly comparing activities in the brains of men and women within a single study.30 Healthy adult subjects (11 women, 11 men) were scanned for regional cerebral glucose utilization while watching a series of emotionally arousing film clips and again while watching a series of matched but more emotionally neutral clips. Memory for the films was assessed in a surprise free recall test three weeks later.
The results showed that a large area of right but not left hemisphere amygdala activity was significantly related to enhanced memory for the emotional film clips in men. An identical analysis in women revealed a large area of left and not right hemisphere amygdala activity related to enhanced memory for the emotional films. The general finding of Cahill et al.30 was confirmed in a separate study of amygdala function employing fMRI.31 Subjects were scanned while viewing a series of emotionally arousing or neutral slides. As reported by Cahill et al.,30 activity of the right and not left amygdala in males related significantly to memory for the most emotional slides. Conversely, activity of the left and not right amygdala related significantly to memory for the most emotional slides in women. Canli et al.31 noted that “both correlations were so robust that they were present even with multiple comparisons across the brain and without selecting the amygdala as a region of interest.”
The single most compelling demonstration of a sex-related hemispheric lateralization to date comes from an fMRI study by Cahill et al.32 Like Canli et al.,31 they used fMRI to examine the relationship of amygdala activity at encoding and subsequent memory for a series of images of varying emotional content. Consistent with the previous studies, activity of the right hemisphere amygdala was significantly more related to subsequent memory for emotional images in men than in women, but activity of the left hemisphere amygdala was significantly more related to subsequent memory for emotional images in women than in men. These findings are shown in Figure 15.2. Crucially, Cahill et al.32 documented a significant interaction between sex and hemisphere in the amygdala relation to memory for emotional material.
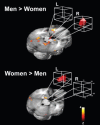
FIGURE 15.2
Sex-related hemispheric lateralization of amygdala function in long-term memory for emotionally arousing films. Activity of the right and not left amygdala in males while viewing emotionally arousing films related significantly to memory of the films (more...)
A simple, but very important conclusion emerges from these studies. No matter what this sex-related lateralization of amygdala function in emotional memory ultimately proves to mean for memory of emotional events in men versus women, these results indicate the studies of the amygdala role in memory (at least for humans) risk conclusions that are incomplete at best and wrong at worst if they fail to account for influences of sex and hemisphere.
15.6. SEX DIFFERENCES IN HUMAN AMYGDALA FUNCTIONAL CONNECTIVITY AT REST
We next examined whether the sex-related hemispheric lateralization of amygdala function in its relation to memory for emotional material might result from differential amygdala connectivity at rest in the absence of emotional stimulation. To do so, we examined the patterns of covariance between the left and right hemisphere amygdalae and the rest of the brain in a large sample of men and women (36 of each sex) who received PET scans while simply resting with their eyes closed.33
The results revealed far wider patterns of covariance between the right hemisphere amygdala and the rest of the brain in men than in women, but far wider patterns of covariance between the left hemisphere and the rest of the brain in women than in men. The key result is shown in Figure 15.3. Although only one slice through the amygdala in the rostral/caudal plane is shown, the pattern was evident throughout the entire extent of the amygdala. All voxels in the right hemisphere displayed significantly greater correlations with the rest of the brain in men than in women, while all voxels in the left hemisphere displayed significantly greater correlations with the rest of the brain in women than in men.

FIGURE 15.3
Location of amygdala seed voxels displaying significant sex-related differences in amygdala functional connectivity during resting conditions. Red areas are associated with greater functional connectivity in women than in men. Blue areas are associated (more...)
An additional, and unsuspected, result concerned the regions in which the amygdalae covaried in the two sexes. In men but not women, the amygdala (right side) covaried with several brain regions (such as the caudate nucleus and visual cortex) important for interactions with the external environment. In women but not men, the amygdala (left side) covaried with several regions (such as the hypothalamus, insular cortex, and cingulate cortex) strongly associated with sensations from the internal milieu. This result raises the intriguing possibility that prior to any overt stimulation there exists in the amygdala at rest a differential “tilt” of its functions toward the external environment in men and toward the internal environment in women.33 This possibility would seem to warrant future investigation.
15.7. POTENTIAL RELATIONSHIP OF SEX-RELATED AMYGDALA HEMISPHERIC SPECIALIZATION TO HEMISPHERIC GLOBAL AND LOCAL PROCESSING BIAS
To better understand implications of the sex-related hemispheric lateralization of amygdala function in relation to explicit memory of emotional events, we integrated it with an already well investigated framework of hemispheric functional specialization positing that the cerebral hemispheres possess differential biases in the processing of relatively global, diffuse versus local, precise aspects of a stimulus or scene.
Substantial evidence from studies of both brain damaged and healthy subjects indicates that the right hemisphere is biased toward the processing of more global holistic aspects of a stimulus or scene, while the left hemisphere is biased towards more local, finer detail processing of the same stimulus or scene.34–40
Considering the evidence of a gender-related hemispheric asymmetry of amygdala function in memory for emotional material described above (males right, females left) and the evidence of hemispheric biases in processing global versus local information (holistic right, detail left), we created a specific hypothesis about a sex-related difference in the effects of a ß-adrenergic blockade on emotional memory. Combining these two lines of evidence with two additional assumptions, namely that amygdala modulatory influences on brain function are predominantly ipsilateral (because amygdalo-cortical projections are almost exclusively ipsilateral41,42 and the amygdala’s modulatory ability requires ß-adrenergic receptor activation43), we predicted that propranolol should impair long-term memory for relatively global (central, gist) aspects of an emotionally arousing story in men, but not memory for relatively local (peripheral detail) aspects of the story. In women, propranolol should produce the opposite effect: impairing memory for peripheral story details but not memory for central story information.
We tested this hypothesis through a re-analysis of published data from two studies demonstrating an impairing effect of ß-adrenergic blockade on memory for an emotionally arousing story.44 Data from the retention tests from these two studies were pooled to increase statistical power and re-analyzed with respect to (1) whether subjects were male or female and (2) whether the questions pertained to central story information or peripheral story detail. Central information was defined as “any information that cannot be changed or removed without changing the fundamental story line,” as determined by consensus of three fourths of independent judges. Figure 15.4 shows the results of this analysis.44

FIGURE 15.4
Recognition test scores for three-phase emotional story. (A) Values for questions defined as pertaining to central information. (B) Values for questions defined as pertaining to peripheral detail. Values represent mean percent correct (± SEM) (more...)
Note in particular the results for story phase 2 (P2 on the x-axis) in which the emotional story elements were introduced (concerning severe injuries to a small boy in an accident while his mother watched) and for which the hypothesis at issue most clearly holds. The P2 results reveal a double dissociation of gender and type of to-be-remembered information (central versus peripheral) on propranolol’s impairing effect on memory: Propranolol significantly impaired P2 memory of central information in men but not women, yet impaired P2 memory of peripheral detail in women but not men.
These results are consistent with the hypothesis that, under emotionally arousing conditions, activation of right amygdala/hemisphere function produces a relative enhancement of memory for central information in males. Activation of left amygdala/hemisphere function in females produces a relative enhancement of memory for peripheral details in women.
15.8. SOME IMPLICATIONS FOR DISEASE STATES
Sex differences exist in the incidence and/or nature of a host of brain-related disorders such as Alzheimer’s disease, schizophrenia, and autism, to name only three.13 More relevant to the present discussion, sex differences also exist in the incidence of emotion-related disorders such as anxiety disorders, post-traumatic stress disorder (PTSD), and clinical depression.
It is well known that PTSD incidence is approximately twice as high in women compared to men.45 Despite this fact, basic science investigations of emotion and memory have left the issue of sex-related influences virtually unexplored. Thus at present we have very little insight into why men and women differ in their susceptibility to PTSD; consequently we know virtually nothing about how we might tailor PTSD treatments to optimally benefit women versus men. Investigations of sex-related influences on emotion and memory should be crucial in helping to fill these important gaps.
This research is also relevant to our understanding of clinical depression, another major disorder about twice as likely to occur in women as in men.46 Interestingly, depression is associated with enhanced activity of the left amygdala in studies predominantly involving women.47 Thus interesting parallels involving amygdala function already exist between mechanisms of emotionally influenced memory in healthy women and clinical depression. It seems likely that studies of the influences of sex on basic mechanisms of emotion and memory will help elucidate neural mechanisms underlying sex differences in depression incidence and potential treatments for depression. Logically, we cannot fully understand disorders with clear sex differences in their incidence and/or nature without careful attention to the influence of sex in our basic science investigations relevant to the disorders.
15.9. SUMMARY
Substantial evidence from animal studies indicates that endogenous stress hormones and the amygdala interact to modulate memory consolidation for emotional events.2 Evidence from human subject studies has clearly reinforced this view. At the same time, it revealed previously unsuspected influences of subject sex on these memory-modulating mechanisms. This evidence combined with a growing number of indications that sex powerfully influences memory-related brain processes13 suggests that future studies of the neurobiology of emotional memory must actively account for the influences of sex. In so doing, the field will be better positioned to understand and treat clinical disorders with sex differences in their incidences and nature.
ACKNOWLEDGMENT
This study was supported by National Institute of Mental Health Grant NIMH RO1 57508.
REFERENCES
- 1.
- Young MP, Scannell JW. Analysis of connectivity: neural systems in the cerebral cortex. Rev Neurosci. 1994;5:227. [PubMed: 7889215]
- 2.
- McGaugh JL. The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annu Rev Neurosci. 2004;27:1. [PubMed: 15217324]
- 3.
- Alkire M, Vazdarjanova A, Dickinson-Anson H, White NS, Cahill L. Selective basolateral amygdala lesions block propofol-induced amnesia. Anesthesiology. 2001;95:708. [PubMed: 11575545]
- 4.
- Cahill L, McGaugh JL. Amygdaloid complex lesions differentially affect retention of tasks using appetitive and aversive reinforcement. Behav Neurosci. 1990;104:532. [PubMed: 2206424]
- 5.
- Small DM, et al. Dissociation of neural representation of intensity and affective valuation in human gustation. Neuron. 2003;39:701. [PubMed: 12925283]
- 6.
- Anderson AK, et al. Dissociated neural representations of intensity and valence in human olfaction. Nat Neurosci. 2003;6:96. [PubMed: 12536208]
- 7.
- Kensinger EA, Corkin S. Two routes to emotional memory: distinct neural processes for valence and arousal. Proc Natl Acad Sci USA. 2004;101:3310. [PMC free article: PMC365786] [PubMed: 14981255]
- 8.
- Lewis PA, Critchley HD, Rothstein P, Dolan RJ. Neural correlates of processing valence and arousal in affective words. Cereb Cortex. 2006 May 22 (Epub preceded print) [PMC free article: PMC2267931] [PubMed: 16699082]
- 9.
- Cahill L, et al. Amygdala activity at encoding correlated with long-term, free recall of emotional information. Proc Natl Acad Sci USA. 1996;93:8016. [PMC free article: PMC38867] [PubMed: 8755595]
- 10.
- Canli T, Desmond JE, Zhao Z, Glover G, Gabrieli JD. Hemispheric asymmetry for emotional stimuli detected with fMRI. Neuroreport. 1998;9:3233. [PubMed: 9831457]
- 11.
- Canli T, Zhao Z, Brewer J, Gabrieli JD, Cahill L. Event-related activation in the human amygdala associates with later memory for individual emotional experience. J Neurosci. 2000;20:RC99. [PMC free article: PMC6772800] [PubMed: 11000199]
- 12.
- Hamann SB, Ely TD, Grafton ST, Kilts CD. Amygdala activity related to enhanced memory for pleasant and aversive stimuli. Nat Neurosci. 2001;2:289. [PubMed: 10195224]
- 13.
- Cahill L. Why sex matters for neuroscience. Nat Rev Neurosci. 2006;7:477. [PubMed: 16688123]
- 14.
- Goldstein JM, et al. Normal sexual dimorphism of the adult human brain assessed by in vivo magnetic resonance imaging. Cereb Cortex. 11:490. [PubMed: 11375910]
- 15.
- George MS, Ketter TA, Parekh PI, Herscovitch P, Post RM. Gender differences in regional cerebral blood flow during transient self-induced sadness or happiness. Biol Psychiatry. 1996;40:859. [PubMed: 8896772]
- 16.
- Killgore WD, Oki M, Yurgelun-Todd DA. Sex-specific developmental changes in amygdala responses to affective faces. Neuroreport. 2001;12:427. [PubMed: 11209962]
- 17.
- Killgore WD, Yurgelun-Todd DA. Sex differences in amygdala activation during the perception of facial affect. Neuroreport. 2001;12:2543. [PubMed: 11496145]
- 18.
- Speck O, Ernst T, Braun J, Koch C, Miller E, Chang L. Gender differences in the functional organization of the brain for working memory. Neuroreport. 2000;11:2581. [PubMed: 10943726]
- 19.
- Baumann SB, Rogers RL, Guinto FC, Saydjari CL, Papanicolaou AC, Eisenberg HM. Gender differences in source location for the N100 auditory evoked magnetic field. Electroencephalogr Clin Neurophysiol. 1991;80:53. [PubMed: 1703950]
- 20.
- Reite M, Sheeder J, Teale P, Richardson D, Adams M, Simon J. MEG based brain laterality: sex differences in normal adults. Neuropsychologia. 1995;33:1607. [PubMed: 8745118]
- 21.
- Salmelin R, et al. Native language, gender, and functional organization of the auditory cortex. Proc Natl Acad Sci USA. 1999;96:10460. [PMC free article: PMC17911] [PubMed: 10468631]
- 22.
- Pugh KR, et al. Cerebral organization of component processes in reading. Brain. 1996;119:1221. [PubMed: 8813285]
- 23.
- Shaywitz BA, et al. Sex differences in the functional organization of the brain for language. Nature. 1995;373:607. [PubMed: 7854416]
- 24.
- Cowan RL, et al. Sex differences in response to red and blue light in human primary visual cortex: a bold fMRI study. Psychiatry Res. 2000;100:129. [PubMed: 11120440]
- 25.
- Li H, et al. Sex differences in cell death. Ann Neurol. 2005;58:317. [PubMed: 15988750]
- 26.
- Wolf OT, Schommer NC, Hellhammer DH, McEwen BS, Kirschbaum C. The relationship between stress induced cortisol levels and memory differs between men and women. Psychoneuroendocrinology. 2001;26:711. [PubMed: 11500252]
- 27.
- Rosvold H, Mirsky A, Pribram K. Influence of amygdalectomy on social behavior in monkeys. J Comp Physiol Psychol. 1954;47:173. [PubMed: 13163250]
- 28.
- Kling A. Differential effects of amygdalectomy in male and female nonhuman primates. Arch Sex Behav. 1974;3:129. [PubMed: 4204411]
- 29.
- Kluver H, Bucy PC. Preliminary analysis of functions of the temporal lobes in monkeys. J Neuropsychiatr Clin Neurosci. 1937;9:606. [PubMed: 9447506]
- 30.
- Cahill L, et al. Sex-related difference in amygdala activity during emotionally influenced memory storage. Neurobiol Learn Mem. 2001;75:1. [PubMed: 11124043]
- 31.
- Canli T, Desmond JE, Zhao Z, Gabrieli JD. Sex differences in the neural basis of emotional memories. Proc Natl Acad Sci USA. 2002;99:10789. [PMC free article: PMC125046] [PubMed: 12145327]
- 32.
- Cahill L, et al. Sex-related hemispheric lateralization of amygdala function in emotionally influenced memory: an fMRI investigation. Learn Mem. 2004;11:261. [PMC free article: PMC419728] [PubMed: 15169855]
- 33.
- Kilpatrick LA, Zald DH, Pardo JV, Cahill LF. Sex-related differences in amygdala functional connectivity during resting conditions. Neuroimage. 2006;30:452. [PubMed: 16326115]
- 34.
- Beeman MJ, Bowden EM. The right hemisphere maintains solution-related activation for yet-to-be-solved problems. Mem Cognit. 2000;28:1231. [PubMed: 11126944]
- 35.
- Delis DC, Robertson LC, Efron R. Hemispheric specialization of memory for visual hierarchical stimuli. Neuropsychologia. 1986;24:205. [PubMed: 3714025]
- 36.
- Fink GR, et al. Where in the brain does visual attention select the forest and the trees? Nature. 1996;382:626. [PubMed: 8757132]
- 37.
- Fink GR, et al. Neural mechanisms involved in the processing of global and local aspects of hierarchically organized visual stimuli. Brain. 1997;120:1779. [PubMed: 9365370]
- 38.
- Fink GR, Marshall JC, Halligan PW, Dolan RJ. Hemispheric asymmetries in global/local processing are modulated by perceptual salience. Neuropsychologia. 1999;37:31. [PubMed: 9920469]
- 39.
- Ivry RB, Robertson LC. The Two Sides of Perception. Bradford; London: 1998.
- 40.
- Sergent J. The cerebral balance of power: confrontation or cooperation? J Exp Psychol Hum Percept Perform. 1982;8:253. [PubMed: 6461721]
- 41.
- Amaral DG, Price JL. Amygdalo-cortical projections in the monkey (Macaca fascicularis). J Comp Neurol. 1984;230:465. [PubMed: 6520247]
- 42.
- Porrino LJ, Crane AM, Goldman-Rakic PS. Direct and indirect pathways from the amygdala to the frontal lobe in rhesus monkeys. J Comp Neurol. 1981;198:121. [PubMed: 6164704]
- 43.
- McGaugh JL, Cahill L, Roozendaal B. Involvement of the amygdala in memory storage: interaction with other brain systems. Proc Natl Acad Sci USA. 1996;93:13508. [PMC free article: PMC33638] [PubMed: 8942964]
- 44.
- Cahill L, van Stegeren A. Sex-related impairment of memory for emotional events with ß-adrenergic blockade. Neurobiol Learn Mem. 2003;79:81. [PubMed: 12482682]
- 45.
- Breslau N, et al. Sex differences in post-traumatic stress disorder. Arch Gen Psychiatr. 1997;54:1044. [PubMed: 9366662]
- 46.
- Kendler KS, Thornton LM, Prescott CA. Gender differences in the rates of exposure to stressful life events and sensitivity to their depressogenic effects. Am J Psychiatr. 2001;158:587. [PubMed: 11282693]
- 47.
- Drevets WC, et al. Glucose metabolism in the amygdala in depression: relationship to diagnostic subtype and plasma cortisol levels. Pharmacol Biochem Behav. 2002;71:431. [PubMed: 11830178]
- Abstract
- INTRODUCTION
- THE AMYGDALA: BUILT TO MODULATE
- HUMAN SUBJECT STUDIES RELATING AMYGDALA ACTIVITY TO EMOTIONAL MEMORY
- AN ASIDE ON SEX DIFFERENCES IN THE BRAIN
- SEX-RELATED INFLUENCES ON AMYGDALA RELATION TO MEMORY FOR EMOTIONAL EVENTS
- SEX DIFFERENCES IN HUMAN AMYGDALA FUNCTIONAL CONNECTIVITY AT REST
- POTENTIAL RELATIONSHIP OF SEX-RELATED AMYGDALA HEMISPHERIC SPECIALIZATION TO HEMISPHERIC GLOBAL AND LOCAL PROCESSING BIAS
- SOME IMPLICATIONS FOR DISEASE STATES
- SUMMARY
- ACKNOWLEDGMENT
- REFERENCES
- Human Brain Imaging Studies of Emotional Memory: Uncovering Influences of Sex an...Human Brain Imaging Studies of Emotional Memory: Uncovering Influences of Sex and Hemisphere - Neural Plasticity and Memory
Your browsing activity is empty.
Activity recording is turned off.
See more...