

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Feingold KR, Anawalt B, Blackman MR, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-.

ABSTRACT
Menstruation is the cyclic, orderly sloughing of the uterine lining, in response to the interactions of hormones produced by the hypothalamus, pituitary, and ovaries. The menstrual cycle may be divided into two phases: (1) follicular or proliferative phase, and (2) the luteal or secretory phase. The length of a menstrual cycle is the number of days between the first day of menstrual bleeding of one cycle to the onset of menses of the next cycle. The median duration of a menstrual cycle is 28 days with most cycle lengths between 25 to 30 days (1-3. Patients who experience menstrual cycles that occur at intervals less than 21 days are termed polymenorrheic, while patients who experience prolonged menstrual cycles greater than 35 days, are termed oligomenorrheic. The typical volume of blood lost during menstruation is approximately 30 mL (4). Any amount greater than 80 mL is considered abnormal (4). The menstrual cycle is typically most irregular around the extremes of reproductive life (menarche and menopause) due to anovulation and inadequate follicular development (5-7). The luteal phase of the cycle is relatively constant in all women, with a duration of 14 days. The variability of cycle length is usually derived from varying lengths of the follicular phase of the cycle, which can range from 10 to 16 days. For complete coverage of this and related topics, please visit www.endotext.org.
THE FOLLICULAR PHASE (Fig.1)
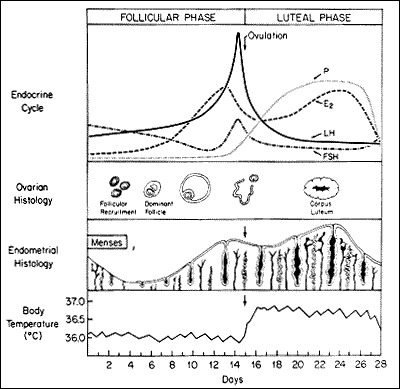
Figure 1.
Hormonal, Ovarian, endometrial, and basal body temperature changes and relations throughout the normal menstrual cycle.
(From Carr BR, Wilson JD. Disorders of the ovary and female reproductive tract. In: Braunwald E, Isselbacher KJ, Petersdorf RG, et al, eds. Harrison's Principles of Internal Medicine. 11th ed. New York: McGraw-Hill, 1987: 1818-1837.
The follicular phase begins from the first day of menses until ovulation. Lower temperatures on a basal body temperature chart, and more importantly, the development of ovarian follicles, characterize this phase. Folliculogenesis begins during the last few days of the preceding menstrual cycle until the release of the mature follicle at ovulation.
Declining steroid production by the corpus luteum and the dramatic fall of inhibin A allows for follicle stimulating hormone (FSH) to rise during the last few days of the menstrual cycle (Fig. 2) (8). Another influential factor on the FSH level in the late luteal phase is related to an increase in GnRH pulsatile secretion secondary to a decline in both estradiol and progesterone levels (9). This elevation in FSH allows for the recruitment of a cohort of ovarian follicles in each ovary, one of which is destined to ovulate during the next menstrual cycle. Once menses ensues, FSH levels begin to decline due to the negative feedback of estrogen and the negative effects of inhibin B produced by the developing follicle (Fig. 2) (8, 10-12). FSH activates the aromatase enzyme in granulosa cells, which converts androgens to estrogen. A decline in FSH levels leads to the production of a more androgenic microenvironment within adjacent follicles to the growing dominant follicle. Also, the granulosa cells of the growing follicle secrete a variety of peptides that may play an autocrine/paracrine role in the inhibition of development of the adjacent follicles.
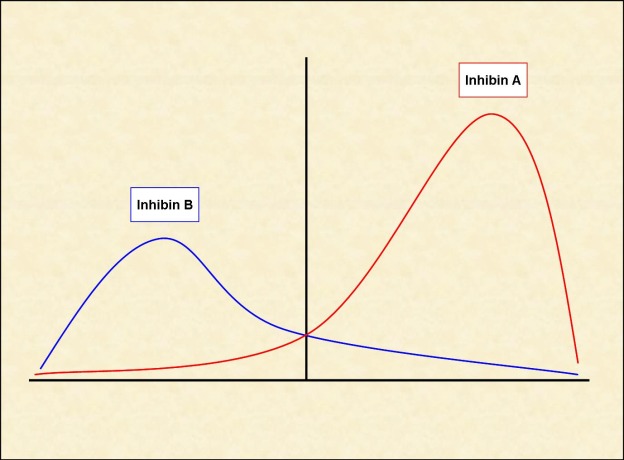
Figure 2.
Inhibin level changes throughout the menstrual cycle.
Inhibin B dominates the follicular phase of the cycle, while Inhibin A dominates the luteal phase.
Development of the dominant follicle has been described in three stages: (1) Recruitment, (2) Selection, and (3) Dominance (Fig.3). The recruitment stage takes place during days 1 through 4 of the menstrual cycle. During this stage, FSH leads to recruitment of a cohort of follicles from the pool of non-proliferating follicles. Between cycle days 5 and 7, selection of a follicle takes place whereby only one follicle is selected from the cohort of recruited follicles to ovulate, and the remaining follicles will undergo atresia. Anti-Müllerian hormone (AMH), a product of granulosa cells, is believed to play a role in the selection of the dominant follicle (13, 14). By cycle day 8, one follicle exerts its dominance by promoting its own growth and suppressing the maturation of the other ovarian follicles thus becoming the dominant follicle.
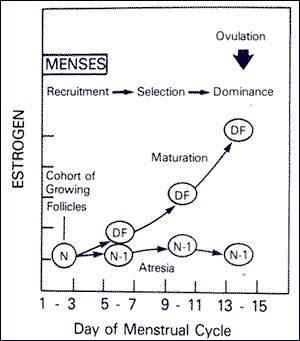
Figure 3.
Time course for recruitment, selection, and ovulation of the dominant ovarian follicle (DF) with onset of atresia among other follicles of the cohort (N-1).
(From Hodgen GD. The dominant ovarian follicle. Fertil Steril 1982; 38:281-300).
During the follicular phase, serum estradiol levels rise in parallel to the growth of follicle size as well as to the increasing number of granulosa cells. FSH receptors exist exclusively on the granulosa cell membranes. Increasing FSH levels during the late luteal phase leads to an increase in the number of FSH receptors and ultimately to an increase in estradiol secretion by granulosa cells. It is important to note that the increase in FSH receptor numbers is due to an increase in the population of granulosa cells and not due to an increase in the concentration of FSH receptors per granulosa cell. Each granulosa cell has approximately 1500 FSH receptors by the secondary stage of follicular development and FSH receptor numbers remains relatively constant for the remainder of development (15). The rise in estradiol secretion appears to increase the total number of estradiol receptors on the granulosa cells (16). In the presence of estradiol, FSH stimulates the formation of LH receptors on granulosa cells allowing for the secretion of small quantities of progesterone and 17-hydroxyprogesterone (17-OHP) which may exert a positive feedback on the estrogen- primed pituitary to augment luteinizing hormone (LH) release (17). FSH also stimulates several steroidogenic enzymes including aromatase, and 3β-hydroxysteroid dehydrogenase (3β-HSD) (18, 19). In table 1, the production rates of sex steroids during the follicular phase, luteal phase, and at the time of ovulation are presented.
Table 1.
Production Rate of Sex Steroids in Women at Different Stages of the Menstrual Cycle
| DAILY PRODUCTION RATE | |||
| SEX STEROIDS* |
Early Follicular | Preovulatory | Mid-luteal |
| Progesterone (mg) | 1 | 4 | 25 |
| 17α-Hydroxyprogesterone (mg) | 0.5 | 4 | 4 |
| Dehydroepiandrosterone (mg) | 7 | 7 | 7 |
| Androstenedione (mg) | 2.6 | 4.7 | 3.4 |
| Testosterone (µg) | 144 | 171 | 126 |
| Estrone (µg) | 50 | 350 | 250 |
| Estradiol (µg) | 36 | 380 | 250 |
From Baird DT. Fraser IS. Blood production and ovarian secretion rates of esuadiol-17β and estrone in women throughout the menstrual cycle. J Clin Endocrinol Metab 38: l009-1017. 1974. @ The Endocrine Society.
- *
Values are expressed in milligrams or micrograms per 24 hours.
In contrast to granulosa cells, LH receptors are located on theca cells during all stages of the menstrual cycle. LH principally stimulates androstenedione production, and to a lesser degree testosterone production in theca cells. In the human, androstenedione is then transported to the granulosa cells where it is aromatized to estrone and finally converted to estradiol by 17-β-hydroxysteroid dehydrogenase type I. This is known as the two-cell, two-gonadotropin hypothesis of regulation of estrogen synthesis in the human ovary (Fig. 4).
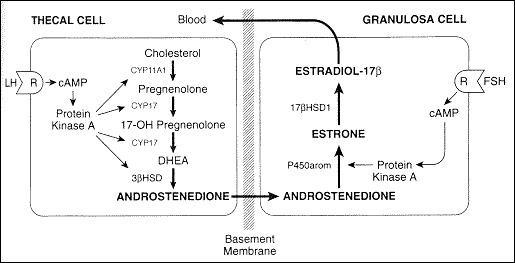
Figure 4.
Two-cell, two-gonadotropin hypothesis of regulation of estrogen synthesis in the human ovary.
Adapted by Carr, BR. Diseases of the ovary and Reproductive Tract. In Wilson JD, Foster DW, Kronenberg HM, Larsen PR, eds. Williams Textbook of Endocrinology 9th edition. WB Saunders, Philadelphia, p.751-817.
In the ovary, the primordial follicles are surrounded by a single layer of granulosa cells and are arrested in the diplotene stage of the first meiotic division. After puberty, each primordial follicle enlarges and develops into a preantral follicle. The preantral follicle is now surrounded by several layers of granulosa cells as well as by theca cells. The preantral follicle is the first stage of FSH receptivity, as now the follicle has acquired FSH receptors. The preantral follicle then develops a cavity and is now known as an antral follicle. Finally, it becomes a preovulatory follicle on its way towards ovulation. Due to the presence of 5α-reductase, preantral and early antral follicles produce more androstenedione and testosterone in relation to estrogens (20). 5α-reductase is the enzyme responsible for converting testosterone to dihydrotestosterone (DHT). Once testosterone has been 5α-reduced, DHT cannot be aromatized. However, the dominant follicle is able to secrete large quantities of estrogen, primarily estradiol, due to high levels of CYP19 (aromatase). This shift from an androgenic to an estrogenic follicular microenvironment may play an important role in selection of the dominant follicle from those follicles that will become atretic.
As mentioned earlier, development of the follicle to the preantral stage is gonadotropin independent, and any follicular growth beyond this point will require gonadotropin interaction. Gonadotropin secretion is regulated by gonadotropin releasing hormone (GnRH), steroid hormones, and various peptides released by the dominant follicle. Also, as mentioned earlier, FSH is elevated during the early follicular phase and then begins to decline until ovulation. In contrast, LH is low during the early follicular phase and begins to rise by the mid-follicular phase due to the positive feedback from the rising estrogen levels. For the positive feedback effect of LH release to occur, estradiol levels must be greater than 200 pg/mL for approximately 50 hours in duration (21). Gonadotropins are normally secreted in a pulsatile fashion from the anterior pituitary, and the frequency and amplitude of the pulses vary according to the phase of the menstrual cycle (Table 2). During the early follicular phase, LH secretion occurs at a pulse frequency of 60 to 90 minutes with relatively constant pulse amplitude. During the late follicular phase prior to ovulation, the pulse frequency increases and the amplitude may begin to increase. In most women, the LH pulse amplitude begins to increase after ovulation takes place (22).
Table 2.
Mean (SEM) Luteinizing Hormone Secretory Burst Characteristics During Phases of the Menstrual Cycle*
|
NUMBER (24 hr) | PERODICITY (min) | AMPLITUDE** (mlU/ml/min) | HALF-DURATIONS (min) | LH HALF-LIFE (min) | TOTAL DAILY SECRETION (mlU/ml/24 hr) |
| Early follicular 175±1.4a | 80 ± 3a | 0.43 ± 0.02a | 6.5 ± 1.0a | 131 ± 13a | 49 ± 6a |
| Late follicular 26.9±1.6b | 53 ± 1b | 0.70 ± 0.03b | 3.5 ± 0.9b | 128 ± 12a | 56 ± 8a |
| Midluteal 10.1±1.0c | 177 ± 15# | 0.26 ± 0.02c# | 11.0 ± 1.1e | 103 ± 7a | 52 ± 4a |
| 395 ± 37d# | 0.95 ± 0.05d# |
- *
Entries in each column identified by a, b, c, d differ significantly (Duncan's multiple-range test, P <.05). Periodicity is intersecretory burst interval. LH, Luteinizing hormone.
**Duration of the deconvolution-resolved LH secretory burst at half-maximal amplitude.
#Maximal rate of LH secretion attained with the deconvolution-resolved LH secretory burst. The midluteal phase has been divided into small (less than 0.65 mIU/ml/min) and large (greater than 0.65 mIU/ml/min) secretory burst amplitudes.Data from Sollenberger MJ, Carlsen EC, Johnson ML, et al. Specific physiological regulation of LH secretory events throughout the human menstrual cycle. New insights into the pulsatile mode of gonadotropin release. J Neuroendocrinol 2:845, 1990.
There are numerous substances found in follicular fluid, such as steroids, pituitary hormones, plasma proteins, proteoglycans and non-steroidal ovarian factors, which regulate the microenvironment of the ovary and regulate steroidogenesis in granulosa cells. Growth factors such as insulin-like growth factor 1 and 2 (IGF1, IGF2) and epidermal growth factor (EGF) are recognized as playing important roles in oocyte development and maturation (23-25). The concentration of ovarian steroids is much higher in follicular fluid in comparison to plasma concentrations. There are 2 populations of antral follicles: (1) large follicles, which are greater than 8mm in diameter, and (2) small follicles, which are less than 8mm. In the large follicles, the concentrations of FSH, estrogen, and progesterone are high while prolactin concentration is low. In the small follicles, prolactin and androgen levels are higher compared to large antral follicles (26).
OVULATION
Ovulation occurs approximately 10-12 hours after the LH peak (Fig. 5) (27). The LH surge is initiated by a dramatic rise of estradiol produced by the preovulatory follicle (Fig. 6). To produce the critical concentration of estradiol needed to initiate the positive feedback, the dominant follicle is almost always >15mm in diameter on ultrasound (28). The beginning of the LH surge occurs roughly 34 to 36 hours prior to ovulation and is a relatively precise predictor for timing ovulation (Fig. 5) (29). The LH surge stimulates luteinization of the granulosa cells and stimulates the synthesis of progesterone responsible for the midcycle FSH surge. Also, the LH surge stimulates resumption of meiosis and the completion of reduction division in the oocyte with the release of the first polar body. It has been demonstrated in cultured granulosa cells that spontaneous luteinization can occur in the absence of LH. It is hypothesized that the inhibitory effects of factors such as oocyte maturation inhibitor or luteinization inhibitor are overcome at ovulation (30).
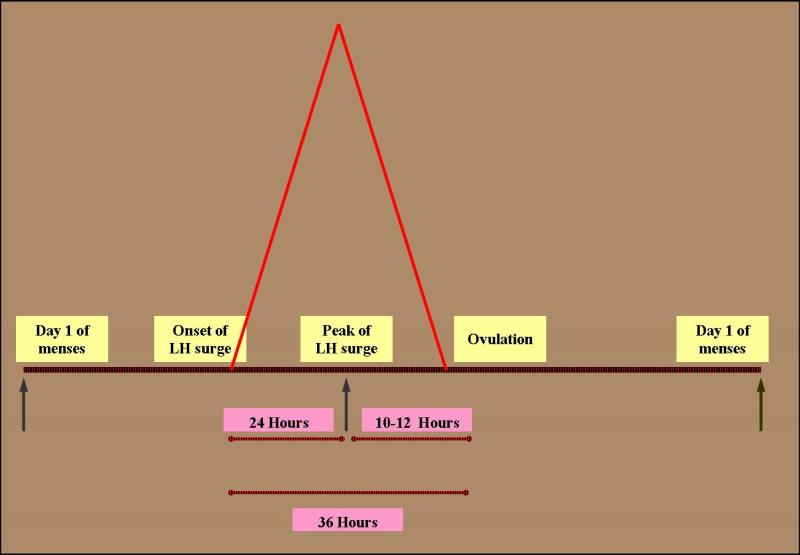
Figure 5.
The onset of LH surge usually precedes ovulation by 36 hours. The peak, on the other hand preceded ovulation by 10-12 hours.

Figure 6.
Changes in gonadotropins and ovarian steroids at midcycle, just prior to ovulation. The initiation of LH surge is at time 0.
Abbreviations: E2, estrogen; P, progesterone (From Hoff JD, Quigley ME, Yen SCC. Hormonal dynamics at midcycle: A re-evaluation. J Clin Endocrinol Metab. 57:792, 1983.
Prostaglandins and proteolytic enzymes, such as collagenase and plasmin, are increased in response to LH and progesterone. Although the precise mechanism is not known, proteolytic enzymes and prostaglandins are activated and digest collagen in the follicular wall, leading to an explosive release of the oocyte-cumulus complex (31). Prostaglandins may also stimulate ovum release by stimulation of smooth muscle within the ovary. The point of the dominant follicle closest to the ovarian surface where this digestion occurs is called the stigma. There is no evidence to support the theory that follicular rupture occurs as a result of increased follicular pressure, although precise measurements precisely at rupture have not been performed (32). In a recent report, laparoscopic visualization of human ovulation during an operative procedure was documented. The authors report visualizing a follicular area called the stigma which was protruding like a bleb from the surface, containing viscous yellow fluid evaginating into the peritoneal cavity (33). In humans, ovulation probably occurs randomly from either ovary during any given cycle. Of interest, some studies have suggested that ovulation occurs more commonly from the right ovary and right sided ovulation carries a higher potential for pregnancy (34). The concentrations of prostaglandins E and F series and hydroxyeicosatetraenoic acid (HETE) reach a peak level in follicular fluid just prior to ovulation (35, 36). Prostaglandins may stimulate proteolytic enzymes while HETEs may stimulate angiogenesis and hyperemia (37). Patients treated with high dose prostaglandin synthetase inhibitors such as Indocin, can have a block in prostaglandin production and effectively block follicular rupture (38-40). This gives rise to what is known as the luteinized, unruptured follicle syndrome and it presents in fertile and infertile patients equally (41). Therefore, infertility patients are advised to avoid taking prostaglandin synthetase inhibitors, as well as cyclo-oxygenase (COX) inhibitors, especially around the time of ovulation (40). A schematic diagram illustrating the proposed mechanisms involved in follicular rupture is presented in Figure 7.
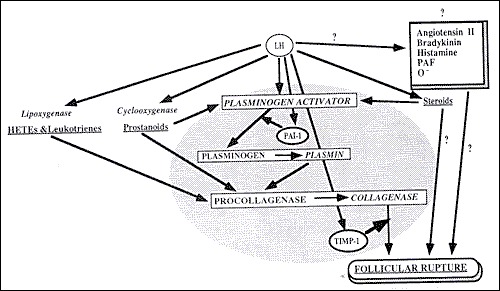
Figure 7.
Proposed mechanisms involved in follicular rupture.
From Tsafriri A, Chun S-Y. Ovulation. In: Adashi E, Rock JA, Rosenwaks Z. Reproductive Endocrinology, Surgery and Technology. Philadelphia: Lippincott-Raven, 1996:236-249.
Estradiol levels fall dramatically immediately prior to the LH peak. This may be due to LH downregulation of its own receptor or because of direct inhibition of estradiol synthesis by progesterone. Progesterone is also responsible for stimulating the midcycle rise in FSH. Elevated FSH levels at this time are thought to free the oocyte from follicular attachments, stimulate plasminogen activator, and increase granulosa cell LH receptors. The mechanism causing the postovulatory fall in LH is unknown. The decline in LH may be due to the loss of the positive feedback effect of estrogen, due to the increasing inhibitory feedback effect of progesterone, or due to a depletion of LH content of the pituitary from downregulation of GnRH receptors (42).
LUTEAL PHASE
This phase is usually 14 days long in most women. After ovulation, the remaining granulosa cells that are not released with the oocyte continue to enlarge, become vacuolated in appearance, and begin to accumulate a yellow pigment called lutein. The luteinized granulosa cells combine with the newly formed theca-lutein cells and surrounding stroma in the ovary to become what is known as the corpus luteum. The corpus luteum is a transient endocrine organ that predominantly secretes progesterone, and its primary function is to prepare the estrogen primed endometrium for implantation of the fertilized ovum. The basal lamina dissolves and capillaries invade into the granulosa layer of cells in response to secretion of angiogenic factors by the granulosa and thecal cells (43). Eight or nine days after ovulation, approximately around the time of expected implantation, peak vascularization is achieved. Figure 8 demonstrates a corpus luteum as seen on transvaginal ultrasound. Note the increased blood flow seen surrounding the corpus luteum as seen with Doppler evaluation. This time also corresponds to peak serum levels of progesterone and estradiol. The central cavity of the corpus luteum may also accumulate with blood and become a hemorrhagic corpus luteum. The life span of the corpus luteum depends upon continued LH support. Corpus luteum function declines by the end of the luteal phase unless human chorionic gonadotropin is produced by a pregnancy. If pregnancy does not occur, the corpus luteum undergoes luteolysis under the influence of estradiol and prostaglandins and forms a scar tissue called the corpus albicans.
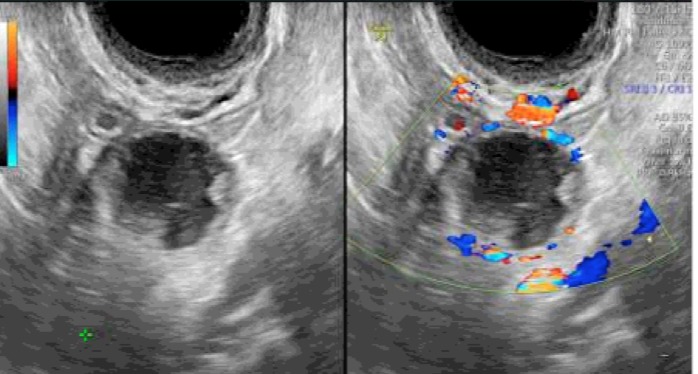
Figure 8.
Corpus luteum as seen on transvaginal ultrasound. On the right image, note the Doppler flow indicating vascular flow surrounding the structure.
Estrogen levels rise and fall twice during the menstrual cycle. Estrogen levels rise during the mid-follicular phase and then drop precipitously after ovulation. This is followed by a secondary rise in estrogen levels during the mid-luteal phase with a decrease at the end of the menstrual cycle. The secondary rise in estradiol parallels the rise of serum progesterone and 17α-hydroxyprogesterone levels. Ovarian vein studies confirm that the corpus luteum is the site of steroid production during the luteal phase (44).
The mechanism by which the corpus luteum regulates steroid secretion is not completely understood. Regulation may be determined in part by LH secretory pattern and LH receptors or variations in the levels of the enzymes regulating steroid hormone production, such as 3β-HSD, CYP17, CYP19, or side chain cleavage enzyme. The number of granulosa cells formed during the follicular phase and the amount of readily available LDL cholesterol may also play a role in steroid regulation by the corpus luteum. The luteal cell population consists of at least two cell types, the large and the small cells (45). Small cells are thought to have been derived from thecal cells while the large cells from granulosa cells. The large cells are more active in steroidogenesis and are influenced by various autocrine/paracrine factors such as inhibin, relaxin, and oxytocin (46, 47).
In studies looking into the mechanisms regulating the menstrual cycle, LH was established as the primary luteotropic agent in a cohort of hypophysectomized women (48). After induction of ovulation, the amount of progesterone secreted and the length of the luteal phase is dependent on repeated LH injections. Administration of LH or HCG during the luteal phase can extend corpus luteum function for an additional two weeks (49).
The secretion of progesterone and estradiol during the luteal phase is episodic, and correlates closely with pulses of LH secretion (Fig. 9) (50). The frequency and amplitude of LH secretion during the follicular phase regulates subsequent luteal phase function and is consistent with the regulatory role of LH during the luteal phase (51). Reduced levels of FSH during the follicular phase can lead to a shortened luteal phase and the development of a smaller corpus lutea (52). Also, the life span of the corpus luteum can be reduced by continuous LH administration during the follicular or luteal phase, reduced LH concentration, decreased LH pulse frequency, or decreased LH pulse amplitude (53-55). The role of other luteotropic factors such as prolactin, oxytocin, inhibin and relaxin is still unclear (56, 57).
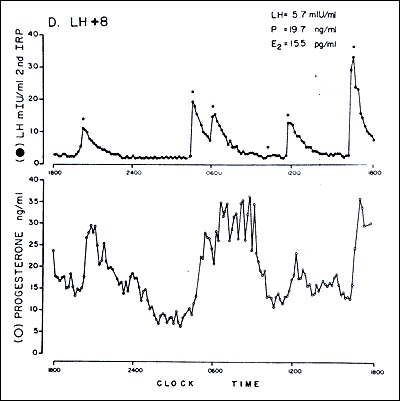
Figure 9.
Episodic secretion of LH (top) and progesterone (bottom) during the luteal phase of a woman.
Abbreviations: LH, luteinizing hormone: P, progesterone E2, estradiol; LH + 8, LH surge plus 8 days. (From Filicori M, Butler JP, Crowley WF Jr. Neuroendocrine regulation of the corpus luteum in the human. J Clin Invest. 73:1638 1984.
The corpus luteum function begins to decline 9-11 days after ovulation. The exact mechanism of how the corpus luteum undergoes its demise is unknown. Estrogen is believed to play a role in the luteolysis of the corpus luteum (58). Estradiol injected into the ovary bearing the corpus luteum induces luteolysis while no effect is noted after estradiol injection of the contralateral ovary (56). However, the absence of estrogen receptors in human luteal cells does not support the role of endogenous estrogen in corpus luteum regression (59). Prostaglandin F2α appears to be luteolytic in nonhuman primates and in studies of women (60, 61). Prostaglandin F2α exerts its effects via the synthesis of endothelin-1, which inhibits steroidogenesis and stimulates the release of a growth factor, tumor necrosis factor alpha (TNFα), which induces cell apoptosis (62). Oxytocin and vasopressin exert their luteotropic effects via an autocrine/paracrine mechanism (63). Luteinizing hormone's ability to downregulate its own receptor may also play a role in termination of the luteal phase. Finally, Matrix metalloproteinases also appear to play a role in luteolysis (64).
Not all hormones undergo marked fluctuations during the normal menstrual cycle. Androgens, glucocorticoids, and pituitary hormones, excluding LH and FSH, undergo only minimal fluctuation (65-68). Due to extra-adrenal 21-hyroxylation of progesterone, plasma levels of deoxycorticosterone are increased during the luteal phase (69, 70).
HORMONAL EFFECTS ON THE REPRODUCTIVE TRACT
Endometrium
The effects of varying concentrations of estrogen and progesterone throughout the course of the menstrual cycle have characteristic effects on the endometrium (Fig. 10) (71). The endometrial changes that occur can be visualized with sonography (Fig. 11). The characteristic endometrial changes also allow for histologic dating. Histologic dating is most accurately accomplished by performing an endometrial biopsy 2-3 days prior to expected menstruation. The proliferative phase is more difficult to date accurately in comparison to the luteal phase. The glands during the proliferative phase are narrow, tubular, and some mitosis and pseudostratification is present. The endometrium thickness is usually between 0.5 and 5mm. In a classical 28-day menstrual cycle, ovulation occurs on day 14. On cycle day 16, the glands take on a more pseudostratified appearance with glycogen accumulating at the basal portion of the glandular epithelium and some nuclei are displaced to the midportion of the cells. In a formalin fixed specimen, glycogen is solubilized resulting in the characteristic basal vacuolization at the base of the endometrial cells. This finding confirms the formation of a functional, progesterone producing, corpus luteum. In the luteal phase, progesterone decreases the biologic activity of estradiol on the endometrium by: (1) decreasing the concentration of estradiol receptors, (2) increasing the enzymatic activity of 17β-hydroxysteroid dehydrogenase type II, the enzyme responsible for the conversion of estradiol to estrone, and (3) by increasing the activity of estrone sulfotransferase (72, 73).
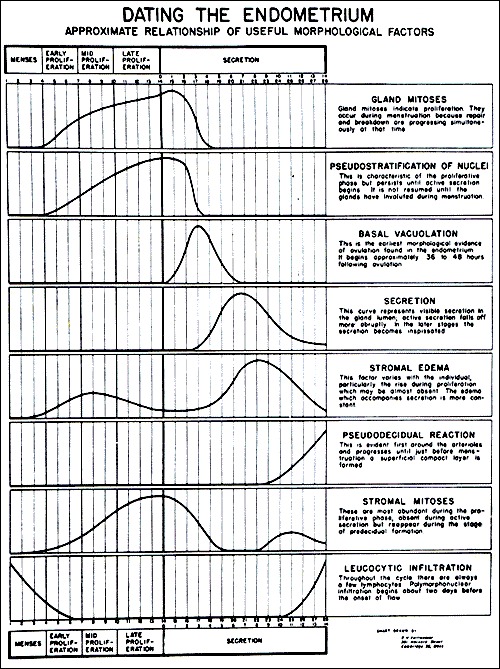
Figure 10.
Dating of the Endometrium.
From Noyes RW, Hertig AW, Rock J. Dating the endometrial biopsy. Fertil Steril 1950; 1:3.
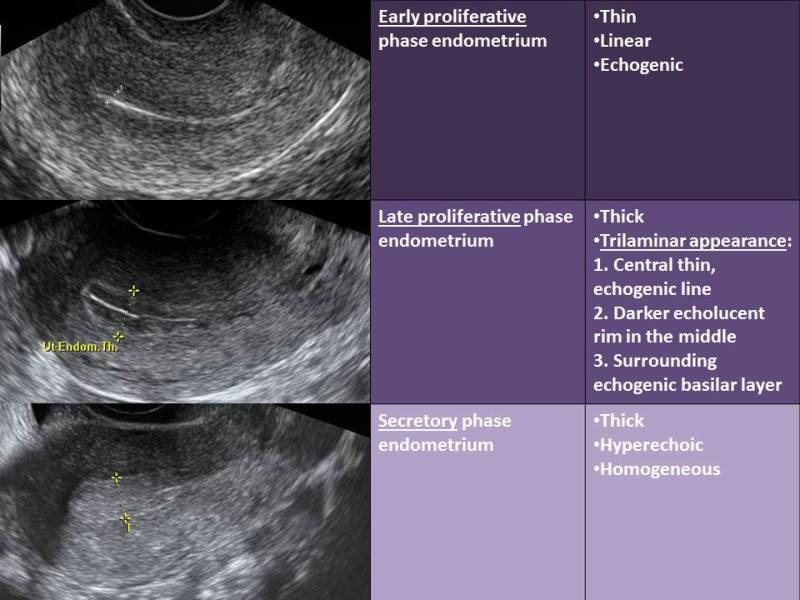
Figure 11.
Characteristic sonographic endometrial changes seen throughout the menstrual cycle.
On cycle day 17, the endometrial glands become more tortuous and dilated. On cycle day 18, the vacuoles in the epithelium decrease in size and are frequently located next to the nuclei. Also, glycogen is now found at the apex of the endometrial cells. By cycle day 19, the pseudostratification and vacuolation almost completely disappear and intraluminal secretions become present. On cycle day 21 or 22, the endometrial stroma begins to become edematous. On cycle day 23, stromal cells surrounding the spiral arterioles begin to enlarge and stromal mitoses become apparent. On cycle day 24, predecidual cells appear around the spiral arterioles and stromal mitoses become more apparent. On cycle day 25, the predecidua begins to differentiate under the surface epithelium. On cycle day 27, there is a marked lymphocytic infiltration and the upper endometrial stroma appears as a solid sheet of well-developed decidua-like cells. On cycle day 28, menstruation begins.
In 2004, Chan et al., were the first to confirm that stem cells were present in human endometrium (74). Subsequent research has involved characterization of the different types of endometrial stem cells (75). Importantly, menstrual fluid may be an easily accessible source for certain types of endometrial stem cells (76). This may lead to advancements in the treatment of many gynecologic disorders including endometriosis and Asherman syndrome as well as non-gynecologic disorders such as neurologic and cardiac disorders (75).
Cervix
The mucous secreting glands of the endocervix are affected by the changes in steroid hormone concentration. Immediately after menstruation, the cervical mucous is scant and viscous. During the late follicular phase, under the influence of rising estradiol levels, the cervical mucous becomes clear, copious and elastic. The quantity of cervical mucous increases 30 fold compared to the early follicular phase (77). The stretchability or elasticity of the cervical mucous can be evaluated between two glass slides and recorded as the spinnbarkeit. If examined under the microscope, the cervical mucous will display a characteristic ferning or palm-leaf arborization appearance. After ovulation, as progesterone levels rise, the cervical mucous once again becomes thick, viscous and opaque and the quantity produced by the endocervical cells decreases.
Vagina
The changes in hormonal levels of estrogen and progesterone also have characteristic effects on the vaginal epithelium. During the early follicular phase, exfoliated vaginal epithelial cells have vesicular nuclei and are basophilic. During the late follicular phase, and the influence of the rising estradiol level, the vaginal epithelial cells display pyknotic nuclei and are acidophilic (78). As progesterone rises during the luteal phase, the acidophilic cells decrease in number and are replaced by an increasing number of leukocytes.
MENSTRUATION
In the absence of a pregnancy, steroid hormone levels begin to fall due to declining corpus luteum function. Progesterone withdrawal results in increased coiling and constriction of the spiral arterioles. This eventually results in tissue ischemia due to decreased blood flow to the superficial endometrial layers, the spongiosa and compacta. The endometrium releases prostaglandins that cause contractions of the uterine smooth muscle and sloughing of the degraded endometrial tissue. The release of prostaglandins may be due to decreased stability of lysosomal membranes in the endometrial cells (79). Infusions of prostaglandin F2α in women during the luteal phase has been shown to induce endometrial necrosis and bleeding (80). The use of prostaglandin synthetase inhibitors decreases the amount of menstrual bleeding and can be used as therapy in women with excessive menstrual bleeding or menorrhagia. Menstrual fluid is composed of desquamated endometrial tissue, red blood cells, inflammatory exudates, and proteolytic enzymes. Within two days after the start of menstruation and while endometrial shedding is still occurring, estrogen produced by the growing follicles starts to stimulate the regeneration of the surface endometrial epithelium. The estrogen secreted by the growing ovarian follicles, causes prolonged vasoconstriction enabling the formation of a clot over the denuded endometrial vessels (81). Also, the regeneration and remodeling of the uterine connective tissue is regulated in part by the matrix metalloproteinase (MMP) system (82).
The average duration of menstrual flow is between four to six days, but the normal range in women can be from as little as two days up to eight days. As mentioned earlier, the average amount of menstrual blood loss is 30 mL and greater than 80 mL is considered abnormal [4].
MENSTRUAL DISORDERS
Apart from conditions of abnormal menstruation, certain disorders are increased in women when compared to men. These conditions are thought to be related to hormone differences as well as hormone changes throughout the menstrual cycle. Increased autoimmune conditions, such as rheumatoid arthritis or systemic lupus erythematosus, are believed to be related to estrogen enhancement of humoral immunity (83). Other researchers also describe higher vulnerability for drug abuse during phases of the menstrual cycle when estradiol levels are high (84).
SUMMARY
The length of a menstrual cycle is the number of days between the first day of menstrual bleeding of one cycle to the onset of menses of the next cycle. The median duration of a menstrual cycle is 28 days with most cycle lengths between 25 to 30 days. The menstrual cycle may be divided into two phases: (1) follicular or proliferative phase, and (2) the luteal or secretory phase. The follicular phase begins from the first day of menses until ovulation. The development of ovarian follicles characterizes this phase. The LH surge is initiated by a dramatic rise of estradiol produced by the preovulatory follicle and results in subsequent ovulation. The LH surge stimulates luteinization of the granulosa cells and stimulates the synthesis of progesterone responsible for the midcycle FSH surge. Also, the LH surge stimulates resumption of meiosis and the completion of reduction division in the oocyte with the release of the first polar body. The luteal phase is 14 days long in most women. If the corpus luteum is not rescued by pregnancy, it will undergo atresia. The resultant progesterone withdrawal results in menses. The average duration of menstrual flow is between four and six days, but the normal range in women can be from as little as two days up to eight days. The average amount of menstrual blood is 30ml, and over 60 ml is considered abnormal.
REFERENCES
- 1.
- Treloar A.E., et al. Variation of the human menstrual cycle through reproductive life. Int J Fertil. 1967;12(1 Pt 2):77–126. [PubMed: 5419031]
- 2.
- Vollman R.F., The Menstrual Cycle. 1977, WB Saunders: Philadelphia. [PubMed: 836520]
- 3.
- Presser, H.B., Temporal data relating to the human menstrual cycle, in Biorhythms and Human Reproduction, M. Ferin, et al., Editors. 1974, John Wiley and Sons: New York. p. 145-160.
- 4.
- Hallberg L., et al. Menstrual blood loss--a population study. Variation at different ages and attempts to define normality. Acta Obstet Gynecol Scand. 1966;45(3):320–351. [PubMed: 5922481]
- 5.
- Apter D., et al. Follicular growth in relation to serum hormonal patterns in adolescent compared with adult menstrual cycles. Fertil Steril. 1987;47(1):82–88. [PubMed: 3098586]
- 6.
- Fraser I.S., et al. Pituitary gonadotropins and ovarian function in adolescent dysfunctional uterine bleeding. J Clin Endocrinol Metab. 1973;37(3):407–414. [PubMed: 4785428]
- 7.
- Lenton E.A., et al. Normal variation in the length of the follicular phase of the menstrual cycle: effect of chronological age. Br J Obstet Gynaecol. 1984;91(7):681–684. [PubMed: 6743609]
- 8.
- Groome N.P., et al. Measurement of dimeric inhibin B throughout the human menstrual cycle. J Clin Endocrinol Metab. 1996;81(4):1401–1405. [PubMed: 8636341]
- 9.
- Welt C.K., et al. Control of follicle-stimulating hormone by estradiol and the inhibins: critical role of estradiol at the hypothalamus during the luteal-follicular transition. J Clin Endocrinol Metab. 2003;88(4):1766–1771. [PubMed: 12679471]
- 10.
- Tsafriri, A., Local nonsteroidal regulators of ovarian function, in The physiology of reproduction, E. Knobil, J.D. Neill, and et al., Editors. 1994, Raven: New York. p. 817-860.
- 11.
- Sawetawan C., et al. Inhibin and activin differentially regulate androgen production and 17 alpha-hydroxylase expression in human ovarian thecal-like tumor cells. J Endocrinol. 1996;148(2):213–221. [PubMed: 8699135]
- 12.
- Welt C.K., et al. Frequency modulation of follicle-stimulating hormone (FSH) during the luteal-follicular transition: evidence for FSH control of inhibin B in normal women. J Clin Endocrinol Metab. 1997;82(8):2645–2652. [PubMed: 9253348]
- 13.
- Durlinger A.L., et al. Control of primordial follicle recruitment by anti-Mullerian hormone in the mouse ovary. Endocrinology. 1999;140(12):5789–96. [PubMed: 10579345]
- 14.
- Hampl R., Snajderova M., Mardesic T. Antimullerian hormone (AMH) not only a marker for prediction of ovarian reserve. Physiol Res. 2011;60(2):217–23. [PubMed: 21114374]
- 15.
- Amsterdam A., Rotmensch S. Structure-function relationships during granulosa cell differentiation. Endocr.Rev. 1987;8(3):309–337. [PubMed: 2820706]
- 16.
- Nimrod A., Erickson G.F., Ryan K.J. A specific FSH receptor in rat granulosa cells: properties of binding in vitro. Endocrinology. 1976;98(1):56–64. [PubMed: 174895]
- 17.
- Fink, G., Gonadotropin secretion and its control, in The physiology of reproduction, E. Knobil, J.D. Neill, and et al., Editors. 1988, Raven: New York. p. 1349-1377.
- 18.
- Zeleznik A.J., Midgley A.R. Jr, Reichert L.E. Jr. Granulosa cell maturation in the rat: increased binding of human chorionic gonadotropin following treatment with follicle-stimulating hormone in vivo. Endocrinology. 1974;95(3):818–825. [PubMed: 4368756]
- 19.
- Erickson G.F., Wang C., Hsueh A.J. FSH induction of functional LH receptors in granulosa cells cultured in a chemically defined medium. Nature. 1979;279(5711):336–338. [PubMed: 221826]
- 20.
- McNatty K.P., et al. Metabolism of androstenedione by human ovarian tissues in vitro with particular reference to reductase and aromatase activity. Steroids. 1979;34(4):429–443. [PubMed: 516112]
- 21.
- Young J.R., Jaffe R.B. Strength-duration characteristics of estrogen effects on gonadotropin response to gonadotropin-releasing hormone in women. II. Effects of varying concentrations of estradiol. J Clin Endocrinol Metab. 1976;42(3):432–442. [PubMed: 767352]
- 22.
- Reame N., et al. Pulsatile gonadotropin secretion during the human menstrual cycle: evidence for altered frequency of gonadotropin-releasing hormone secretion. J Clin Endocrinol Metab. 1984;59(2):328–337. [PubMed: 6429184]
- 23.
- Zhou J., Bondy C. Anatomy of the human ovarian insulin-like growth factor system. Biol Reprod. 1993;48(3):467–482. [PubMed: 7680905]
- 24.
- Maruo T., et al. Expression of epidermal growth factor and its receptor in the human ovary during follicular growth and regression. Endocrinology. 1993;132(2):924–931. [PubMed: 8425504]
- 25.
- Thierry van Dessel H.J., et al. Serum and follicular fluid levels of insulin-like growth factor I (IGF-I), IGF-II, and IGF-binding protein-1 and -3 during the normal menstrual cycle. J Clin Endocrinol Metab. 1996;81(3):1224–31. [PubMed: 8772603]
- 26.
- Hillier S.G., et al. Intraovarian sex steroid hormone interactions and the regulation of follicular maturation: aromatization of androgens by human granulosa cells in vitro. J Clin Endocrinol Metab. 1980;50(4):640–647. [PubMed: 6767736]
- 27.
- Pauerstein C.J., et al. Temporal relationships of estrogen, progesterone, and luteinizing hormone levels to ovulation in women and infrahuman primates. Am J Obstet Gynecol. 1978;130(8):876–886. [PubMed: 416719]
- 28.
- Cahill D.J., et al. Onset of the preovulatory luteinizing hormone surge: diurnal timing and critical follicular prerequisites. Fertil Steril. 1998;70(1):56–59. [PubMed: 9660421]
- 29.
- Hoff J.D., Quigley M.E., Yen S.S. Hormonal dynamics at midcycle: a reevaluation. J Clin Endocrinol Metab. 1983;57(4):792–796. [PubMed: 6411753]
- 30.
- Channing, C.P., et al., Ovarian follicular and luteal physiology, in International Review of Physiology, R.O. Greep, Editor. 1980, University Park Press: Baltimore. p. 117. [PubMed: 6248477]
- 31.
- Espey, L.L. and H. Lipner, Ovulation, in The Physiology of Reproduction, E. Knobil and J.D. Neill, Editors. 1994, Raven: New York. p. 725.
- 32.
- Espey L.L. Ovarian proteolytic enzymes and ovulation. Biol Reprod. 1974;10(2):216–235. [PubMed: 4462824]
- 33.
- Lousse J.C., Donnez J. Laparoscopic observation of spontaneous human ovulation. Fertil Steril. 2008;90(3):833–834. [PubMed: 18440526]
- 34.
- Fukuda M., et al. Right-sided ovulation favours pregnancy more than left-sided ovulation. Hum Reprod. 2000;15(9):1921–1926. [PubMed: 10966987]
- 35.
- Lumsden M.A., et al. Changes in the concentration of prostaglandins in preovulatory human follicles after administration of hCG. J Reprod Fertil. 1986;77(1):119–124. [PubMed: 3459888]
- 36.
- Espey L.L., et al. Ovarian hydroxyeicosatetraenoic acids compared with prostanoids and steroids during ovulation in rats. Am J Physiol. 1991;260(2 Pt 1):E163–E169. [PubMed: 1996618]
- 37.
- Sherman B.M., West J.H., Korenman S.G. The menopausal transition: analysis of LH, FSH, estradiol, and progesterone concentrations during menstrual cycles of older women. J Clin Endocrinol Metab. 1976;42(4):629–636. [PubMed: 1262439]
- 38.
- O'Grady J.P., et al. The effects of an inhibitor of prostaglandin synthesis (indomethacin) on ovulation, pregnancy, and pseudopregnancy in the rabbit. Prostaglandins. 1972;1(2):97–106. [PubMed: 4660072]
- 39.
- Killick S., Elstein M. Pharmacologic production of luteinized unruptured follicles by prostaglandin synthetase inhibitors. Fertil Steril. 1987;47(5):773–777. [PubMed: 3552753]
- 40.
- Pall M., Friden B.E., Brannstrom M. Induction of delayed follicular rupture in the human by the selective COX-2 inhibitor rofecoxib: a randomized double-blind study. Hum Reprod. 2001;16(7):1323–1328. [PubMed: 11425807]
- 41.
- Doody, K.J. and B.R. Carr, Diagnosis and treatment of luteal dysfunction, in Ovarian Endocrinology, S.G. Hillier, Editor. 1991, Blackwell Scientific: London. p. 260.
- 42.
- Katt J.A., et al. The frequency of gonadotropin-releasing hormone stimulation determines the number of pituitary gonadotropin-releasing hormone receptors. Endocrinology. 1985;116(5):2113–2115. [PubMed: 2985372]
- 43.
- Koos R.D. Potential relevance of angiogenic factors to ovarian physiology. Semin Reprod Endocrinol. 1989;7:29.
- 44.
- Niswender, G.D. and T.M. Nett, The corpus luteum and its control in infraprimate species, in The Physiology of Reproduction, E. Knobil and J.D. Neill, Editors. 1994, Raven: New York. p. 781.
- 45.
- Retamales I., et al. Morpho-functional study of human luteal cell subpopulations. Hum Reprod. 1994;9(4):591–596. [PubMed: 8046008]
- 46.
- Khan-Dawood F.S., et al. Human corpus luteum secretion of relaxin, oxytocin, and progesterone. J Clin Endocrinol Metab. 1989;68(3):627–631. [PubMed: 2918060]
- 47.
- Carr B.R., et al. The effect of transforming growth factor-beta on steroidogenesis and expression of key steroidogenic enzymes with a human ovarian thecal-like tumor cell model. Am J Obstet Gynecol. 1996;174(4):1109–1116. [PubMed: 8623837]
- 48.
- Vande Wiele R.L., et al. Mechanisms regulating the menstrual cycle in women. Recent.Prog.Horm.Res. 1970;26:63–103. [PubMed: 4919096]
- 49.
- Segaloff A., Sternberg W.H., Gaskill C.J. Effects of luteotropic dose of chorionic gonadotropin in women. J Clin Endocrinol Metab. 1951;11(9):936–944. [PubMed: 14873766]
- 50.
- Filicori M., Butler J.P., Crowley W.F. Jr. Neuroendocrine regulation of the corpus luteum in the human. Evidence for pulsatile progesterone secretion. J Clin Invest. 1984;73(6):1638–1647. [PMC free article: PMC437074] [PubMed: 6427277]
- 51.
- McNeely M.J., Soules M.R. The diagnosis of luteal phase deficiency: a critical review. Fertil Steril. 1988;50(1):1–15. [PubMed: 3289975]
- 52.
- Stouffer R.L., Hodgen G.D. Induction of luteal phase defects in rhesus monkeys by follicular fluid administration at the onset of the menstrual cycle. J Clin Endocrinol Metab. 1980;51(3):669–671. [PubMed: 6773981]
- 53.
- Sheehan K.L., Casper R.F., Yen S.S. Luteal phase defects induced by an agonist of luteinizing hormone-releasing factor: a model for fertility control. Science. 1982;215(4529):170–172. [PubMed: 6797068]
- 54.
- Keyes P.L., Wiltbank M.C. Endocrine regulation of the corpus luteum. Annu.Rev Physiol. 1988;50:465–482. [PubMed: 3288101]
- 55.
- Cooke I.D. The corpus luteum. Hum Reprod. 1988;3(2):153–156. [PubMed: 3281962]
- 56.
- Auletta F.J., Flint A.P. Mechanisms controlling corpus luteum function in sheep, cows, nonhuman primates, and women especially in relation to the time of luteolysis. Endocr.Rev. 1988;9(1):88–105. [PubMed: 3286237]
- 57.
- Hodgen G.D. Surrogate embryo transfer combined with estrogen-progesterone therapy in monkeys. Implantation, gestation, and delivery without ovaries. JAMA. 1983;250(16):2167–2171. [PubMed: 6620521]
- 58.
- Gore B.Z., Caldwell B.V., Speroff L. Estrogen-induced human luteolysis. J Clin Endocrinol Metab. 1973;36(3):615–617. [PubMed: 4685397]
- 59.
- Iwai T., et al. Immunohistochemical localization of oestrogen receptors and progesterone receptors in the human ovary throughout the menstrual cycle. Virchows Arch A Pathol Anat Histopathol. 1990;417(5):369–375. [PubMed: 2122584]
- 60.
- Wentz A.C., Jones G.S. Transient luteolytic effect of prostaglandin F2alpha in the human. Obstet Gynecol. 1973;42(2):172–181. [PubMed: 4721404]
- 61.
- Schoonmaker J.N., et al. Estradiol-induced luteal regression in the rhesus monkey: evidence for an extraovarian site of action. Endocrinology. 1982;110(5):1708–1715. [PubMed: 6804210]
- 62.
- Shikone T., et al. Apoptosis of human corpora lutea during cyclic luteal regression and early pregnancy. J Clin Endocrinol Metab. 1996;81(6):2376–2380. [PubMed: 8964880]
- 63.
- Khan-Dawood F.S., Huang J.C., Dawood M.Y. Baboon corpus luteum oxytocin: an intragonadal peptide modulator of luteal function. Am J Obstet Gynecol. 1988;158(4):882–891. [PubMed: 3364500]
- 64.
- Young K.A., Hennebold J.D., Stouffer R.L. Dynamic expression of mRNAs and proteins for matrix metalloproteinases and their tissue inhibitors in the primate corpus luteum during the menstrual cycle. Mol Hum Reprod. 2002;8(9):833–840. [PubMed: 12200461]
- 65.
- Judd H.L., Yen S.S. Serum androstenedione and testosterone levels during the menstrual cycle. J Clin Endocrinol Metab. 1973;36(3):475–481. [PubMed: 4685386]
- 66.
- Genazzani A.R., et al. Pattern of plasma ACTH, hGH, and cortisol during menstrual cycle. J Clin Endocrinol Metab. 1975;41(3):431–437. [PubMed: 169283]
- 67.
- Carr B.R., et al. Plasma levels of adrenocorticotropin and cortisol in women receiving oral contraceptive steroid treatment. J Clin Endocrinol Metab. 1979;49(3):346–349. [PubMed: 224073]
- 68.
- Carr, B.R. and J.D. Wilson, Disorders of the ovary and the female reproductive tract, in Harrison's Principles of Internal Medicine, E. Braunwald, et al., Editors. 1987, McGraw-Hill: New York. p. 1818.
- 69.
- Casey M.L., MacDonald P.C. Extraadrenal formation of a mineralocorticosteroid: deoxycorticosterone and deoxycorticosterone sulfate biosynthesis and metabolism. Endocr.Rev. 1982;3(4):396–403. [PubMed: 6759106]
- 70.
- Parker C.R. Jr, et al. Plasma concentrations of 11-deoxycorticosterone in women during the menstrual cycle. Obstet Gynecol. 1981;58(1):26–30. [PubMed: 6454093]
- 71.
- Noyes R.W., Hertig A.T., Rock J. Dating the endometrial biopsy. Am J Obstet Gynecol. 1975;122(2):262–263. [PubMed: 1155504]
- 72.
- Lessey B.A., et al. Immunohistochemical analysis of human uterine estrogen and progesterone receptors throughout the menstrual cycle. J Clin Endocrinol Metab. 1988;67(2):334–340. [PubMed: 2455728]
- 73.
- Tseng L., Liu H.C. Stimulation of arylsulfotransferase activity by progestins in human endometrium in vitro. J Clin Endocrinol Metab. 1981;53(2):418–421. [PubMed: 6454696]
- 74.
- Chan RWS, Schwab KE, Gargett CE. Clonogenicity of human endometrial epithelial and stromal cells. Biol Reprod. 2004;70:1738–50. [PubMed: 14766732]
- 75.
- Gargett CE, Schwab KE, Deane JA. Endometrial stem/progenitor cells: the first 10 years. Hum Reprod Update. 2015;22:dmv051. [PMC free article: PMC4755439] [PubMed: 26552890]
- 76.
- Patel AN, Park E, Kuzman M, Benetti F, Silva FJ, Allickson JG: Multipotent menstrual blood stromal stem cells: isolation, characterization, and differentiation. Cell Transplant 2008 [cited 2018 Mar 13];17:303–11. [PubMed: 18522233]
- 77.
- Rebar, R.E., Practical evaluation of hormonal status, in Reproductive Endocrinology: Physiology, Pathophysiology and Clinical Management, S.S.C. Yen and R.B. Jaffe, Editors. 1991, WB Saunders: Philadelphia. p. 830.
- 78.
- Gaudefroy M. Cytologic criteria of estrogen effect. Acta Cytol. 1958;(2):347.
- 79.
- Henzl M.R., et al. Lysosomal concept of menstrual bleeding in humans. J Clin Endocrinol Metab. 1972;34(5):860–875. [PubMed: 5012496]
- 80.
- Turksoy R.N., Safaii H.S. Immediate effect of prostaglandin F2alpha during the luteal phase of the menstrual cycle. Fertil Steril. 1975;26(7):634–637. [PubMed: 168099]
- 81.
- Edman C.D. The effects of steroids on the endometrium. Semin Reprod Endocrinol. 1983;1(3):179.
- 82.
- Curry T.E. Jr, Osteen K.G. Cyclic changes in the matrix metalloproteinase system in the ovary and uterus. Biol Reprod. 2001;64(5):1285–1296. [PubMed: 11319131]
- 83.
- Cutolo M. Gender and the rheumatic diseases: Epidemiological evidence and possible biologic mechanisms. Annals of the Rheumatic Diseases. 2003;62:3–3.
- 84.
- Anker J.J., Carroll M.E. Females are more vulnerable to drug abuse than males: evidence from preclinical studies and the role of ovarian hormones. Curr Top Behav Neurosci. 2011;8:73–96. [PubMed: 21769724]
- Menstrual bleeding, cycle length, and follicular and luteal phase lengths in women without known subfertility: A pooled analysis of three cohorts.[Paediatr Perinat Epidemiol. 2020]Menstrual bleeding, cycle length, and follicular and luteal phase lengths in women without known subfertility: A pooled analysis of three cohorts.Najmabadi S, Schliep KC, Simonsen SE, Porucznik CA, Egger MJ, Stanford JB. Paediatr Perinat Epidemiol. 2020 May; 34(3):318-327. Epub 2020 Feb 27.
- Characteristics of menstrual cycles with or without intercourse in women with no known subfertility.[Hum Reprod Open. 2022]Characteristics of menstrual cycles with or without intercourse in women with no known subfertility.Najmabadi S, Schliep KC, Simonsen SE, Porucznik CA, Egger MJ, Stanford JB. Hum Reprod Open. 2022; 2022(4):hoac039. Epub 2022 Sep 27.
- Onapristone (ZK 98.299): a potential antiprogestin for endometrial contraception.[Am J Obstet Gynecol. 1995]Onapristone (ZK 98.299): a potential antiprogestin for endometrial contraception.Katkam RR, Gopalkrishnan K, Chwalisz K, Schillinger E, Puri CP. Am J Obstet Gynecol. 1995 Sep; 173(3 Pt 1):779-87.
- Review Menstrual Cycle Changes as Women Approach the Final Menses: What Matters?[Obstet Gynecol Clin North Am. ...]Review Menstrual Cycle Changes as Women Approach the Final Menses: What Matters?Harlow SD. Obstet Gynecol Clin North Am. 2018 Dec; 45(4):599-611. Epub 2018 Oct 25.
- Review Mode of action of RU 486.[Ann Med. 1993]Review Mode of action of RU 486.Bygdeman M, Swahn ML, Gemzell-Danielsson K, Svalander P. Ann Med. 1993 Feb; 25(1):61-4.
- The Normal Menstrual Cycle and the Control of Ovulation - EndotextThe Normal Menstrual Cycle and the Control of Ovulation - Endotext
Your browsing activity is empty.
Activity recording is turned off.
See more...