To form an anchoring junction, cells must first adhere. A bulky cytoskeletal apparatus must then be assembled around the molecules that directly mediate the adhesion. The result is a well-defined structure—a desmosome, a hemidesmosome, a focal adhesion, or an adherens junction—that is easily identified in the electron microscope. Indeed, electron microscopy provided the basis for the original classification of cell junctions. In the early stages of cell junction development, however, before the cytoskeletal apparatus has assembled, cells often adhere to one another without clearly displaying these characteristic structures; in the electron microscope, one may simply see two plasma membranes separated by a small gap of a definite width. Functional tests show, nevertheless, that the two cells are stuck to each other, and biochemical analysis can reveal the molecules responsible for the adhesion.
The study of cell-cell junctions and the study of cell-cell adhesion were once quite distinct endeavors, originating from two different experimental approaches—junctions through electron microscopic description, and adhesion through functional tests and biochemistry. Only in recent years have these two approaches begun to converge in a unified view of the molecular basis of cell junctions and cell adhesion. In the previous section, we concentrated on the structures of mature cell junctions. In this section, we turn to functional and biochemical studies of the cell-cell adhesion mechanisms that operate when cells migrate over other cells and when they assemble into tissues—mechanisms that precede the construction of mature cell-cell anchoring junctions. We begin with a critical question for embryonic development: what mechanisms ensure that a cell attaches to appropriate neighbors at the right time?
Animal Cells Can Assemble into Tissues Either in Place or After They Migrate
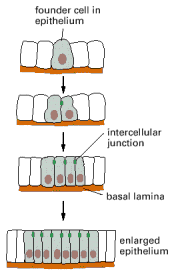
Many simple tissues, including most epithelial tissues, derive from precursor cells whose progeny are prevented from wandering away by being attached to the extracellular matrix, to other cells, or to both (). But the accumulating cells do not simply remain passively stuck together; instead, the tissue architecture is generated and actively maintained by selective adhesions that the cells make and progressively adjust.
The simplest mechanism by which cells assemble to form a tissue. The progeny of the founder cell are retained in the epithelium by the basal lamina and by cell-cell adhesion mechanisms, including the formation of intercellular junctions.
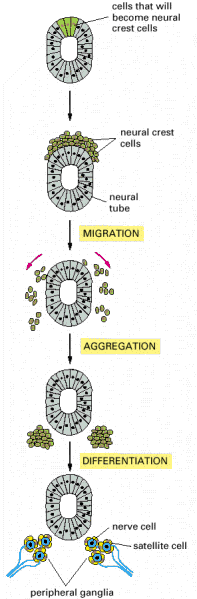
Selective adhesion is even more essential for the development of tissues that have more complex origins involving cell migration. In these tissues, one population of cells invades another and assembles with it, and perhaps with other migrant cells, to form an orderly structure. In vertebrate embryos, for example, cells from the neural crest break away from the epithelial neural tube, of which they are initially a part, and migrate along specific paths to many other regions (discussed in Chapter 21). There they assemble with other cells and with one another to differentiate into a variety of tissues, including those of the peripheral nervous system ().
An example of a more complex mechanism by which cells assemble to form a tissue. Some cells that are initially part of the epithelial neural tube alter their adhesive properties and disengage from the epithelium to form the neural crest on the upper surface (more...)
Cell motility and cell adhesion combine to bring about these kinds of morphogenetic events. The process requires some mechanism for directing the cells to their final destination. This may involve chemotaxis or chemorepulsion, the secretion of a soluble chemical that attracts or repels migrating cells, respectively, or pathway guidance, the laying down of adhesive or repellent molecules in the extracellular matrix or on cell surfaces to guide the migrating cells along the right paths. Then, once a migrating cell has reached its destination, it must recognize and join other cells of the appropriate type to assemble into a tissue. How this latter process occurs can be studied if cells of different embryonic tissues are artificially mingled, after which they often spontaneously sort out to restore a more normal arrangement, as we discuss next.
Dissociated Vertebrate Cells Can Reassemble into Organized Tissues Through Selective Cell-Cell Adhesion
Unlike adult vertebrate tissues, which are difficult to dissociate, embryonic vertebrate tissues are easily dissociated. This is usually done by treating the tissue with low concentrations of a proteolytic enzyme such as trypsin, sometimes combined with the removal of extracellular Ca2+ and Mg2+ with a divalent-cation chelator (such as EDTA). These reagents disrupt the protein-protein interactions (many of which are divalent-cation-dependent) that hold cells together. Remarkably, the dissociated cells often reassemble in vitro into structures that resemble the original tissue. Such findings reveal that tissue structure is not just a product of history; it is actively maintained and stabilized by the system of affinities that cells have for one another and for the extracellular matrix.
A striking example of this phenomenon is seen when dissociated cells from two embryonic vertebrate organs, such as the liver and the retina, are mixed together and artificially formed into a pellet: the mixed aggregates gradually sort out according to their organ of origin. More generally, disaggregated cells are found to adhere more readily to aggregates of their own organ than to aggregates of other organs. Evidently there are cell-cell recognition systems that make cells of the same differentiated tissue preferentially adhere to one another; these adhesive preferences are presumably important in stabilizing tissue architecture.
Cells adhere to each other and to the extracellular matrix through cell-surface proteins called cell adhesion molecules (CAMs)—a category that includes the transmembrane adhesion proteins we have already discussed. CAMs can be cell-cell adhesion molecules or cell-matrix adhesion molecules. Some CAMs are Ca2+-dependent, whereas others are Ca2+-independent. The Ca2+-dependent CAMs seem to be primarily responsible for the tissue-specific cell-cell adhesion seen in early vertebrate embryos, explaining why these cells can be disaggregated with Ca2+-chelating agents.
CAMs were initially identified by making antibodies against cell-surface molecules and then testing the antibodies for their ability to inhibit cell-cell adhesion in a test tube. Those rare antibodies that inhibit the adhesion were then used to characterize and isolate the adhesion molecule recognized by the antibodies.
Cadherins Mediate Ca2+-dependent Cell-Cell Adhesion
The cadherins are the major CAMs responsible for Ca2+-dependent cell-cell adhesion in vertebrate tissues. The first three cadherins that were discovered were named according to the main tissues in which they were found: E-cadherin is present on many types of epithelial cells; N-cadherin on nerve, muscle, and lens cells; and P-cadherin on cells in the placenta and epidermis. All are also found in various other tissues; N-cadherin, for example, is expressed in fibroblasts, and E-cadherin is expressed in parts of the brain. These and other classical cadherins are related in sequence throughout their extracellular and intracellular domains. There are also a large number of nonclassical cadherins, with more than 50 expressed in the brain alone. The nonclassical cadherins include proteins with known adhesive function, such as the desmosomal cadherins discussed earlier and the diverse protocadherins found in the brain. They also include proteins that appear to have nonadhesive functions, such as T-cadherin, which lacks a transmembrane domain and is attached to the plasma membrane of nerve and muscle cells by a glycosylphosphatidylinositol (GPI) anchor, and the Fat protein, which was first identified as the product of a tumor-suppressor gene in Drosophila. Together, the classical and nonclassical cadherin proteins constitute the cadherin superfamily ().
Some Members of the Cadherin Superfamily.
Cadherins are expressed in both invertebrates and vertebrates. Virtually all vertebrate cells seem to express one or more cadherins, according to the cell type. They are the main adhesion molecules holding cells together in early embryonic tissues. In culture, the removal of extracellular Ca2+ or treatment with anti-cadherin antibodies disrupts embryonic tissues, and, if cadherin-mediated adhesion is left intact, antibodies against other adhesion molecules have little effect. Mutations that inactivate the function of E-cadherin cause mouse embryos to fall apart and die early in development.
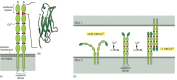
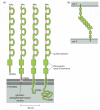
Most cadherins are single-pass transmembrane glycoproteins about 700–750 amino acids long. Structural studies suggest that they associate in the plasma membrane to form dimers or larger oligomers. The large extracellular part of the polypeptide chain is usually folded into five or six cadherin repeats, which are structurally related to immunoglobulin (Ig) domains ( and ). The crystal structures of E- and N-cadherin have helped to explain the importance of Ca2+ binding for cadherin function. The Ca2+ ions are positioned between each pair of cadherin repeats, locking the repeats together to form a stiff, rodlike structure: the more Ca2+ ions that are bound, the more rigid the structure is. If Ca2+ is removed, the extracellular part of the protein becomes floppy and is rapidly degraded by proteolytic enzymes ().
The structure and function of cadherins. (A) A classical cadherin molecule. The protein is a homodimer, with the extracellular part of each polypeptide folded into five cadherin repeats. There are Ca2+-binding sites between each pair of repeats. (B) The (more...)
Cadherins Have Crucial Roles in Development
E-cadherin is the best-characterized cadherin. It is usually concentrated in adherens junctions in mature epithelial cells, where it helps connect the cortical actin cytoskeletons of the cells it holds together (see ). E-cadherin is also the first cadherin expressed during mammalian development. It helps cause compaction, an important morphological change that occurs at the eight-cell stage of mouse embryo development. During compaction, the loosely attached cells, called blastomeres, become tightly packed together and joined by intercellular junctions. Antibodies against E-cadherin block blastomere compaction, whereas antibodies that react with various other cell-surface molecules on these cells do not.
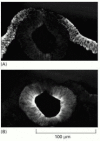
It seems likely that cadherins are also crucial in later stages of vertebrate development, since their appearance and disappearance correlate with major morphogenetic events in which tissues segregate from one another. As the neural tube forms and pinches off from the overlying ectoderm, for example, neural tube cells lose E-cadherin and acquire other cadherins, including N-cadherin, while the cells in the overlying ectoderm continue to express E-cadherin (). Then, when the neural crest cells migrate away from the neural tube, these cadherins become scarcely detectable, and another cadherin (cadherin-7) appears that helps hold the migrating cells together as loosely associated cell groups. Finally, when the cells aggregate to form a ganglion, they re-express N-cadherin (see ).
The distribution of E-cadherin and N-cadherin in the developing nervous system. Immunofluorescence micrographs of a cross section of a chick embryo showing the developing neural tube labeled with antibodies against (A) E-cadherin and (B) N-cadherin. Note (more...)
If N-cadherin is overexpressed in the emerging neural crest cells, the cells fail to escape from the neural tube. Thus, not only do cell groups that originate from one cell layer exhibit distinct patterns of cadherin expression when separating from one another, but these switches in cadherin expression seem to be intimately involved in the separation process.
Cadherins Mediate Cell-Cell Adhesion by a Homophilic Mechanism
How do cell-cell adhesion molecules such as the cadherins bind cells together? Three possibilities are illustrated in : (1) in homophilic binding, molecules on one cell bind to other molecules of the same kind on adjacent cells; (2) in heterophilic binding, the molecules on one cell bind to molecules of a different kind on adjacent cells; (3) in linker-dependent binding, cell-surface receptors on adjacent cells are linked to one another by secreted multivalent linker molecules. Although all three mechanisms have been found to operate in animals, cadherins usually link cells by the homophilic mechanism. In a line of cultured fibroblasts called L cells, for example, the cells neither express cadherins nor adhere to one another. When L cells are transfected with DNA encoding E-cadherin, the transfected cells become adherent to one another by a Ca2+-dependent mechanism, and the adhesion is inhibited by anti-E-cadherin antibodies. Since cadherin proteins can bind directly to one another and the transfected cells do not bind to untransfected L cells, one can conclude that E-cadherin binds cells together through the interaction of two E-cadherin molecules on different cells.
Three mechanisms by which cell-surface molecules can mediate cell-cell adhesion. Although all of these mechanisms can operate in animals, the one that depends on an extracellular linker molecule seems to be the least common.
If L cells expressing different cadherins are mixed together, they sort out and aggregate separately, indicating that different cadherins preferentially bind to their own type (), mimicking what happens when cells derived from tissues that express different cadherins are mixed together. A similar segregation of cells occurs if L cells expressing different amounts of the same cadherin are mixed together (). It therefore seems likely that both qualitative and quantitative differences in the expression of cadherins have a role in organizing tissues.
Cadherin-dependent cell sorting. Cells in culture can sort themselves out according to the type and level of cadherins they express. This can be visualized by labeling different populations of cells with dyes of different colors. (A) Cells expressing (more...)
In the nervous system especially, there are many different cadherins, each with a distinct but overlapping pattern of expression (). As they are concentrated at synapses, they are thought to have a role in synapse formation and stabilization. Some of the nonclassical cadherins, such as the protocadherins, are strong candidates for helping to determine the specificity of synaptic connections. Like antibodies, they differ in their N-terminal (variable) regions but are identical in their C-terminal (constant) regions. The extracellular variable region and intracellular constant region are encoded by separate exons, with the variable-region exons arranged in tandem arrays upstream of the constant-region exons (). The diversity of protocadherins is generated by a combination of differential promoter usage and alternative RNA splicing, rather than by site-specific recombination as occurs in antibody diversification (discussed in Chapter 24).
Cadherin diversity in the central nervous system. (A) Expression patterns for three classical cadherins in the embryonic mouse brain. (B) The arrangement of exons that encode the members of one of the three known protocadherin families of nonclassical (more...)
Cadherins Are Linked to the Actin Cytoskeleton by Catenins
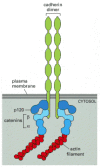
Most cadherins, including all classical and some nonclassical ones, function as transmembrane adhesion proteins that indirectly link the actin cytoskeletons of the cells they join together. This arrangement occurs in adherens junctions (see ). The highly conserved cytoplasmic tail of these cadherins interacts indirectly with actin filaments by means of a group of intracellular anchor proteins called catenins (). This interaction is essential for efficient cell-cell adhesion, as classical cadherins that lack their cytoplasmic domain cannot hold cells strongly together.
The linkage of classical cadherins to actin filaments. The cadherins are coupled indirectly to actin filaments by the anchor proteins α-catenin and β-catenin. A third intracellular protein, called p120, also binds to the cadherin cytoplasmic (more...)
As discussed earlier, the nonclassical cadherins that form desmosomes interact with intermediate filaments, rather than with actin filaments. Their cytoplasmic domain binds to a different set of intracellular anchor proteins, which in turn bind to intermediate filaments (see ).
Some cells can regulate the adhesive activity of their cadherins. This regulation may be important for the cellular rearrangements that occur within epithelia when these cell sheets change their shape and organization during animal development (see ). The molecular basis of this regulation is uncertain but may involve the phosphorylation of anchor proteins attached to the cytoplasmic tail of the cadherins.
Some cadherins can help transmit signals to the cell interior. Vascular endothelial cadherin (VE-cadherin), for example, not only mediates adhesion between endothelial cells but also is required for endothelial cell survival. Although endothelial cells that do not express VE-cadherin still adhere to one another via N-cadherin, they do not survive (see , p. 1082). Their survival depends on an extracellular signal protein called vascular endothelial growth factor (VEGF), which binds to a receptor tyrosine kinase (discussed in Chapter 15) that uses VE-cadherin as a co-receptor.
Selectins Mediate Transient Cell-Cell Adhesions in the Bloodstream
White blood cells lead a nomadic life, moving to and fro between the bloodstream and the tissues, and this necessitates special adhesive properties. These properties depend on selectins. Selectins are cell-surface carbohydrate-binding proteins (lectins) that mediate a variety of transient, Ca2+-dependent, cell-cell adhesion interactions in the bloodstream. There are at least three types: L-selectin on white blood cells, P-selectin on blood platelets and on endothelial cells that have been locally activated by an inflammatory response, and E-selectin on activated endothelial cells. Each selectin is a transmembrane protein with a highly conserved lectin domain that binds to a specific oligosaccharide on another cell ().
The structure and function of selectins. (A) The structure of P-selectin. The selectin attaches to the actin cytoskeleton through anchor proteins that are still poorly characterized. (B) How selectins and integrins mediate the cell-cell adhesions required (more...)
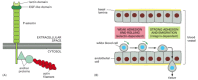
Selectins have an important role in binding white blood cells to endothelial cells lining blood vessels, thereby enabling the blood cells to migrate out of the bloodstream into a tissue. In a lymphoid organ, the endothelial cells express oligosaccharides that are recognized by L-selectin on lymphocytes, causing the lymphocytes to loiter and become trapped. Conversely, at sites of inflammation, the endothelial cells switch on expression of selectins, which recognize the oligosaccharides on white blood cells and platelets, flagging the cells down to help deal with the local emergency. Selectins do not act alone, however; they collaborate with integrins, which strengthen the binding of the blood cells to the endothelium. The cell-cell adhesions mediated by both selectins and integrins are heterophilic (see ): selectins bind to specific oligosaccharides on glycoproteins and glycolipids, while integrins bind to specific proteins.
Selectins and integrins act in sequence to let white blood cells leave the bloodstream and enter tissues. The selectins mediate a weak adhesion because the binding of the lectin domain of the selectin to its carbohydrate ligand is of low affinity. This allows the white blood cell to adhere weakly and reversibly to the endothelium, rolling along the surface of the blood vessel propelled by the flow of blood. The rolling continues until the blood cell activates its integrins (discussed later), now causing the cell to bind strongly to the endothelial cell surface and to crawl out of the blood vessel between adjacent endothelial cells ().
Members of the Immunoglobulin Superfamily of Proteins Mediate Ca2+-independent Cell-Cell Adhesion
Cadherins, selectins, and integrins all depend on extracellular Ca2+ (or Mg2+ for some integrins) to function in cell adhesion. The molecules responsible for Ca2+-independent cell-cell adhesion belong mainly to the large and ancient immunoglobulin (Ig) superfamily of proteins. These proteins contain one or more Ig-like domains that are characteristic of antibody molecules (discussed in Chapter 24). One of the best-studied examples is the neural cell adhesion molecule (N-CAM), which is expressed by a variety of cell types, including most nerve cells. N-CAM is the most prevalent of the Ca2+-independent cell-cell adhesion molecules in vertebrates, and, like cadherins, it is thought to bind cells together by a homophilic mechanism (between N-CAM molecules on adjacent cells). Some Ig-like cell-cell adhesion proteins, however, use a heterophilic mechanism. Intercellular adhesion molecules (ICAMs) on endothelial cells, for example, bind to integrins on blood cells when blood cells migrate out of the bloodstream, as just discussed.
There are at least 20 forms of N-CAM, all generated by alternative splicing of an RNA transcript produced from a single gene. In all forms, the large extracellular part of the polypeptide chain is folded into five Ig-like domains (). Some forms of N-CAM carry an unusually large quantity of sialic acid (with chains containing hundreds of repeating sialic acid units). By virtue of their negative charge, these long polysialic acid chains hinder cell adhesion, and there is increasing evidence that N-CAM heavily loaded with sialic acid serves to prevent adhesion, rather than cause it.
The cell adhesion protein N-CAM. (A) Four forms of N-CAM. The extracellular part of the polypeptide chain in each case is folded into five Ig-like domains (and one or two other domains called fibronectin type III repeats). Disulfide bonds (red) connect (more...)
Although cadherins and Ig family members are frequently expressed on the same cells, the adhesions mediated by cadherins are much stronger, and they are largely responsible for holding cells together, segregating cell collectives into discrete tissues, and maintaining tissue integrity. N-CAM and other members of the Ig family seem to contribute more to the fine-tuning of these adhesive interactions during development and regeneration. In the developing rodent pancreas, for example, the formation of the islets of Langerhans requires cell aggregation, followed by cell sorting. Whereas inhibition of cadherin function prevents cell aggregation and islet formation, loss of N-CAM only impairs the cell sorting process, so that disorganized islets form.
Similarly, whereas mutant mice that lack N-cadherin die early in development, mutant mice that lack N-CAM develop relatively normally, although they do have some defects in neural development. Mutations in other genes that encode Ig-like cell adhesion proteins, however, can cause more severe neural defects. L1 gene mutations in humans, for example, cause mental retardation and other neurological defects resulting from abnormalities in the migration of nerve cells and their axons.
The importance of Ig-like cell adhesion proteins in connecting the neurons of the developing nervous system has been demonstrated dramatically in Drosophila. An N-CAM-like protein called fasciclin III (FAS3) is expressed transiently on some motor neurons, as well as on the muscle cells they normally innervate. If FAS3 is genetically removed from these neurons, they fail to recognize their muscle targets and do not make synapses with them. Conversely, if motor neurons that normally do not express FAS3 are made to express this protein, they now synapse with FAS3-expressing muscle cells to which they normally do not connect. It seems that FAS3 mediates these synaptic connections by a homophilic “matchmaking” mechanism.
Like the cadherins, some Ig-like proteins do more than just bind cells together. They can also transmit signals to the cell interior. Some forms of N-CAM in nerve cells, for example, associate with Src family cytoplasmic tyrosine kinases (discussed in Chapter 15), which relay signals onward by phosphorylating intracellular proteins on tyrosines. Other Ig family members are transmembrane tyrosine phosphatases (discussed in Chapter 15) that help guide growing axons to their target cells, presumably by dephosphorylating specific intracellular proteins.
Multiple Types of Cell-Surface Molecules Act in Parallel to Mediate Selective Cell-Cell Adhesion
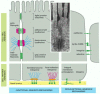
A single type of cell utilizes multiple molecular mechanisms in adhering to other cells. Some of these mechanisms involve organized cell junctions, while others do not (). Each cell in a multicellular animal contains an assortment of cell-surface receptors that enables the cell to respond specifically to a complementary set of soluble extracellular signal molecules, such as hormones and growth factors. Likewise, each cell in a tissue has a particular combination (and concentration) of cell-surface adhesion molecules that enables it to bind in its own characteristic way to other cells and to the extracellular matrix. And just as receptors for soluble extracellular signal molecules generate intracellular signals that alter the cell's behavior, so too can cell adhesion molecules, although the signaling mechanisms they use are generally not as well understood.
A summary of the junctional and nonjunctional adhesive mechanisms used by animal cells in binding to one another and to the extracellular matrix. The junctional mechanisms are shown in epithelial cells, while the nonjunctional mechanisms are shown in (more...)
Unlike receptors for soluble signal molecules, which bind their specific ligand with high affinity, the receptors that bind to molecules on cell surfaces or in the extracellular matrix usually do so with relatively low affinity. These low-affinity receptors rely on the enormous increase in binding strength gained through the simultaneous binding of multiple receptors to multiple ligands on an opposing cell or in the adjacent matrix. One could call this the “Velcro principle.”
We have seen, however, that the interaction of the extracellular binding domains of these cell-surface molecules is not enough to ensure cell adhesion. At least in the case of cadherins and, as we shall see, integrins, the adhesion molecules must also attach (via anchor proteins) to the cytoskeleton inside the cell. The cytoskeleton is thought to assist and stabilize the lateral clustering of the adhesion molecules to facilitate multipoint binding. The cytoskeleton is also required to enable the adhering cell to exert traction on the adjacent cell or matrix (and vice versa). Thus, the mixture of specific types of cell-cell adhesion molecules present on any two cells, as well as their concentration, cytoskeletal linkages, and distribution on the cell surface, determine the total affinity with which the two cells bind to each other.
Nonjunctional Contacts May Initiate Cell-Cell Adhesions That Junctional Contacts Then Orient and Stabilize
We have seen that adhesive contacts between cells play a crucial part in organizing the formation of tissues and organs in developing embryos or in adult tissues undergoing repair after injury. Most often, these contacts do not involve the formation of organized intercellular junctions that show up as specialized structures in the electron microscope. The interacting plasma membranes are simply seen to come close together and run parallel, separated by a space of 10–20 nm. This type of “nonjunctional” contact may be optimal for cell locomotion—close enough to provide traction and to allow transmembrane adhesion proteins to interact, but not so tight, or so solidly anchored to the cytoskeleton, as to immobilize the cell.
A reasonable hypothesis is that nonjunctional cell-cell adhesion proteins initiate cell-cell adhesions, which are then oriented and stabilized by the assembly of full-blown intercellular junctions. Many of the transmembrane proteins involved can diffuse in the plane of the plasma membrane and, in this or other ways, can be recruited to sites of cell-cell (and cell-matrix) contact, enabling nonjunctional adhesions to enlarge and mature into junctional adhesions. This has been demonstrated for some integrins and cadherins, which help initiate cell adhesion and then later become integral parts of cell junctions. The migrating tip of an axon, for example, has an even distribution of cadherins on its surface, which helps it adhere to other cells along the migration pathway. It also has an intracellular pool of cadherins in vesicles just under the plasma membrane. When the axon reaches its target cell, it is thought to release the intracellular cadherin molecules onto the cell surface, where they help form a stable contact, which matures into a chemical synapse.
As discussed earlier, antibodies against adherens junction proteins block the formation of tight junctions, as well as adherens junctions, suggesting that the assembly of one type of junction can be a prerequisite for the formation of another. An increasing number of monoclonal antibodies and peptide fragments have been produced that can block a single type of cell adhesion molecule. Moreover, an increasing number of genes encoding these cell-surface proteins have been identified, creating new opportunities for manipulating the adhesive machinery of cells in culture and in experimental animals. It is now possible, therefore, to inactivate the various cell-cell adhesion proteins in combinations—a requirement for deciphering the rules of cell-cell recognition and binding used to build complex tissues.
Summary
Cells dissociated from various tissues of vertebrate embryos preferentially reassociate with cells from the same tissue when they are mixed together. This tissue-specific recognition process in vertebrates is mediated mainly by a family of Ca2+-dependent cell-cell adhesion proteins called cadherins, which hold cells together by a homophilic interaction between these transmembrane proteins on adjacent cells. For this interaction to be effective, the cytoplasmic part of the cadherins must be linked to the cytoskeleton by cytoplasmic anchor proteins called catenins.
Two other families of transmembrane adhesion proteins have major roles in cell-cell adhesion. Selectins function in transient Ca2+-dependent cell-cell adhesions in the bloodstream by binding to specific oligosaccharides on the surface of another cell. Members of the immunoglobulin superfamily, including N-CAM, mediate Ca2+-independent cell-cell adhesion processes that are especially important during neural development.
Even a single cell type uses multiple molecular mechanisms in adhering to other cells (and to the extracellular matrix). Thus, the specificity of cell-cell (and cell-matrix) adhesion seen in embryonic development must result from the integration of several different adhesion systems, of which some are associated with specialized cell junctions, while others are not.