We normally think of pathogens in hostile terms—as invaders that attack our bodies. But a pathogen or a parasite, like any other organism, is simply trying to live and procreate. Living at the expense of a host organism is a very attractive strategy, and it is possible that every living organism on earth is subject to some type of infection or parasitism (). A human host is a nutrient-rich, warm, and moist environment, which remains at a uniform temperature and constantly renews itself. It is not surprising that many microorganisms have evolved the ability to survive and reproduce in this desirable niche. In this section, we discuss some of the common features that microorganisms must have in order to be infectious. We then explore the wide variety of organisms that are known to cause disease in humans.
Parasitism at many levels. (A) Scanning electron micrograph of a flea. The flea is a common parasite of mammals—including dogs, cats, rats, and humans. It drinks the blood of its host. Flea bites spread bubonic plague by passing the pathogenic (more...)
Pathogens Have Evolved Specific Mechanisms for Interacting with Their Hosts
The human body is a complex and thriving ecosystem. It contains about 1013 human cells and also about 1014 bacterial, fungal, and protozoan cells, which represent thousands of microbial species. These microbes, called the normal flora, are usually limited to certain areas of the body, including the skin, mouth, large intestine, and vagina. In addition, humans are always infected with viruses, most of which rarely, if ever, become symptomatic. If it is normal for us to live in such close intimacy with a wide variety of microbes, how is it that some of them are capable of causing us illness or death?
Pathogens are usually distinct from the normal flora. Our normal microbial inhabitants only cause trouble if our immune systems are weakened or if they gain access to a normally sterile part of the body (for example, when a bowel perforation enables the gut flora to enter the peritoneal cavity of the abdomen, causing peritonitis). In contrast, dedicated pathogens do not require that the host be immunocompromised or injured. They have developed highly specialized mechanisms for crossing cellular and biochemical barriers and for eliciting specific responses from the host organism that contribute to the survival and multiplication of the pathogen.
In order to survive and multiply in a host, a successful pathogen must be able to: (1) colonize the host; (2) find a nutritionally compatible niche in the host body; (3) avoid, subvert, or circumvent the host innate and adaptive immune responses; (4) replicate, using host resources; and (5) exit and spread to a new host. Under severe selective pressure to induce only the correct host cell responses to accomplish this complex set of tasks, pathogens have evolved mechanisms that maximally exploit the biology of their host organisms. Many of the pathogens we discuss in this chapter are skillful and practical cell biologists. We can learn a great deal of cell biology by observing them.
The Signs and Symptoms of Infection May Be Caused by the Pathogen or by the Host's Responses
Although we can easily understand why infectious microorganisms would evolve to reproduce in a host, it is less clear why they would evolve to cause disease. One explanation may be that, in some cases, the pathological responses elicited by microorganisms enhance the efficiency of their spread or propagation and hence clearly have a selective advantage for the pathogen. The virus-containing lesions on the genitalia caused by herpes simplex infection, for example, facilitate direct spread of the virus from an infected host to an uninfected partner during sexual contact. Similarly, diarrheal infections are efficiently spread from patient to caretaker. In many cases, however, the induction of disease has no apparent advantage for the pathogen.
Many of the symptoms and signs that we associate with infectious disease are direct manifestations of the host's immune responses in action. Some hallmarks of bacterial infection, including the swelling and redness at the site of infection and the production of pus (mainly dead white blood cells), are the direct result of immune system cells attempting to destroy the invading microorganisms. Fever, too, is a defensive response, as the increase in body temperature can inhibit the growth of some microorganisms. Thus, understanding the biology of an infectious disease requires an appreciation of the contributions of both pathogen and host.
Pathogens Are Phylogenetically Diverse
Many types of pathogens cause disease in humans. The most familiar are viruses and bacteria. Viruses cause diseases ranging from AIDS and smallpox to the common cold. They are essentially fragments of nucleic acid (DNA or RNA) instructions, wrapped in a protective shell of proteins and (in some cases) membrane (). They use the basic transcription and translation machinery of their host cells for their replication.
Pathogens in many forms. (A) The structure of the protein coat, or capsid, of poliovirus. This virus was once a common cause of paralysis, but the disease (poliomyelitis) has been nearly eradicated by widespread vaccination. (B) The bacterium Vibrio cholerae (more...)
Of all the bacteria we encounter in our lives, only a small minority are dedicated pathogens. Much larger and more complex than viruses, bacteria are usually free-living cells, which perform most of their basic metabolic functions themselves, relying on the host primarily for nutrition ().
Some other infectious agents are eucaryotic organisms. These range from single-celled fungi and protozoa (), through large complex metazoa such as parasitic worms. One of the most common infectious diseases on the planet, shared by about a billion people at present, is an infestation in the gut by Ascaris lumbricoides. This nematode closely resembles its cousin Caenorhabditis elegans, which is widely used as a model organism for genetic and developmental biological research (discussed in Chapter 21). C. elegans, however, is only about 1 mm in length, whereas Ascaris can reach 30 cm ().
Some rare neurodegenerative diseases, including mad cow disease, are caused by an unusual type of infectious particle called a prion, which is made only of protein. Although the prion contains no genome, it can nevertheless replicate and kill the host.
Even within each class of pathogen, there is striking diversity. Viruses vary tremendously in their size, shape, and content (DNA versus RNA, enveloped or not, and so on), and the same is true for the other pathogens. The ability to cause disease (pathogenesis) is a lifestyle choice, not a legacy shared only among close relatives ().
Phylogenetic diversity of pathogens. This diagram shows the similarities among 16S ribosomal RNA for cellular life forms (bacteria, archaea, and eucaryotes). Each branch is labeled with the name of a representative member of that group, and the length (more...)
Each individual pathogen causes disease in a different way, which makes it challenging to understand the basic biology of infection. But, when considering the interactions of infectious agents with their hosts, some common themes of pathogenesis emerge. These common themes are the focus of this chapter. First, we introduce the basic features of each of the major types of pathogens that exploit features of host cell biology. Then, we examine in turn the mechanisms that pathogens use to control their hosts and the innate mechanisms that hosts use to control pathogens.
Bacterial Pathogens Carry Specialized Virulence Genes
Bacteria are small and structurally simple, compared to the vast majority of eucaryotic cells. Most can be classified broadly by their shape as rods, spheres, or spirals and by their cell-surface properties. Although they lack the elaborate morphological variety of eucaryotic cells, they display a surprising array of surface appendages that enable them to swim or to adhere to desirable surfaces (). Their genomes are correspondingly simple, typically on the order of 1,000,000–5,000,000 nucleotide pairs in size (compared to 12,000,000 for yeast and more than 3,000,000,000 for humans).
Bacterial shapes and cell-surface structures. Bacteria are classified into three different shapes: (A) spheres (cocci), (B) rods (bacilli), and (C) spiral cells (spirochetes). (D) They are also classified as Gram-positive or Gram-negative. Bacteria such (more...)
As emphasized above, only a minority of bacterial species have developed the ability to cause disease in humans. Some of those that do cause disease can only replicate inside the cells of the human body and are called obligate pathogens. Others replicate in an environmental reservoir such as water or soil and only cause disease if they happen to encounter a susceptible host; these are called facultative pathogens. Many bacteria are normally benign but have a latent ability to cause disease in an injured or immunocompromised host; these are called opportunistic pathogens.
Some bacterial pathogens are fastidious in their choice of host and will only infect a single species or a group of related species, whereas others are generalists. Shigella flexneri, for example, which causes epidemic dysentery (bloody diarrhea) in areas of the world lacking a clean water supply, will only infect humans and other primates. By contrast, the closely related bacterium Salmonella enterica, which is a common cause of food poisoning in humans, can also infect many other vertebrates, including chickens and turtles. A champion generalist is the opportunistic pathogen Pseudomonas aeruginosa, which is capable of causing disease in plants as well as animals.
The significant differences between a virulent pathogenic bacterium and its closest nonpathogenic relative may result from a very small number of genes. Genes that contribute to the ability of an organism to cause disease are called virulence genes. The proteins they encode are called virulence factors. Virulence genes are frequently clustered together, either in groups on the bacterial chromosome called pathogenicity islands or on extrachromosomal virulence plasmids (). These genes may also be carried on mobile bacteriophages (bacterial viruses). It seems therefore that a pathogen may arise when groups of virulence genes are transferred together into a previously avirulent bacterium. Consider, for example, Vibrio cholerae—the bacterium that causes cholera. Several of the genes encoding the toxins that cause the diarrhea in cholera are carried on a mobile bacteriophage (). Of the hundreds of strains of Vibrio cholerae found in lakes in the wild, the only ones that cause human disease are those that have become infected with this virus.
Genetic differences between pathogens and nonpathogens. Nonpathogenic Escherichia coli has a single circular chromosome. E. coli is very closely related to two types of food-borne pathogens—Shigella flexneri, which causes dysentery, and Salmonella (more...)
Genetic organization of Vibrio cholerae. (A) Vibrio cholerae is unusual in having two circular chromosomes rather than one. The two chromosomes have distinct origins of replication (oriC1 and oriC2). Three loci in pathogenic strains of V. cholerae are (more...)
Many virulence genes encode proteins that interact directly with host cells. Two of the genes carried by the Vibrio cholerae phage, for example, encode two subunits of cholera toxin. The B subunit of this secreted, toxic protein binds to a glycolipid component of the plasma membrane of the epithelial cells in the gut of a person who has consumed Vibrio cholerae in contaminated water. The B subunit transfers the A subunit through the membrane into the epithelial cell cytoplasm. The A subunit is an enzyme that catalyzes the transfer of an ADP-ribose moiety from NAD to the trimeric G protein Gs, which normally activates adenylyl cyclase to make cyclic AMP (discussed in Chapter 15). ADP-ribosylation of the G protein results in an overaccumulation of cyclic AMP and an ion imbalance, leading to the massive watery diarrhea associated with cholera. The infection is then spread by the fecal-oral route by contaminated food and water.
Some pathogenic bacteria use several independent mechanisms to cause toxicity to the cells of their host. Anthrax, for example, is an acute infectious disease of sheep, cattle, other herbivores, and occasionally humans. It is usually caused by contact with spores of the Gram-positive bacterium, Bacillus anthracis. Unlike cholera, anthrax has never been observed to spread directly from one infected person to another. Dormant spores can survive in soil for long periods of time and are highly resistant to adverse environmental conditions, including heat, ultraviolet and ionizing radiation, pressure, and chemical agents. After the spores are inhaled, ingested, or rubbed into breaks in the skin, the spores germinate, and the bacteria begin to replicate. Growing bacteria secrete two toxins, called lethal toxin and edema toxin. Either toxin alone is sufficient to cause signs of infection. Like the A and B subunits of cholera toxin, both toxins are made of two subunits. The B subunit is identical between lethal toxin and edema toxin, and it binds to a host cell-surface receptor to transfer the two different A subunits into host cells. The A subunit of edema toxin is an adenylyl cyclase that directly converts host cell ATP into cyclic AMP. This causes an ion imbalance that can lead to accumulation of extracellular fluid (edema) in the infected skin or lung. The A subunit of lethal toxin is a zinc protease that cleaves several members of the MAP kinase kinase family (discussed in Chapter 15). Injection of lethal toxin into the bloodstream of an animal causes shock and death. The molecular mechanisms and the sequence of events leading to death in anthrax remain uncertain.
These examples illustrate a common theme among virulence factors. They are frequently either toxic proteins (toxins) that directly interact with important host structural or signaling proteins to elicit a host cell response that is beneficial to pathogen colonization or replication, or they are proteins that are needed to deliver such toxins to their host cell targets. One common and particularly efficient delivery mechanism, called the type III secretion system, acts like a tiny syringe that injects toxic proteins from the cytoplasm of an extracellular bacterium directly into the cytoplasm of an adjacent host cell (). There is a remarkable degree of structural similarity between the type III syringe and the base of a bacterial flagellum (see ), and many of the proteins in the two structures are clearly homologous.
Type III secretion systems that can deliver virulence factors into the cytoplasm of host cells. (A) Electron micrographs of purified type III apparatuses. About two dozen proteins are necessary to make the complete structure, which is seen in the three (more...)
Because bacteria form a kingdom distinct from the eucaryotes they infect (see ), much of their basic machinery for DNA replication, transcription, translation, and fundamental metabolism is quite different from that of their host. These differences enable us to find antibacterial drugs that specifically inhibit these processes in bacteria, without disrupting them in the host. Most of the antibiotics that we use to treat bacterial infections are small molecules that inhibit macromolecular synthesis in bacteria by targeting bacterial enzymes that are either distinct from their eucaryotic counterparts or that are involved in pathways, such as cell wall biosynthesis, that are absent in humans ( and Table 6-3).
Antibiotic targets. Despite the large number of antibiotics available, they have a narrow range of targets, which are highlighted in yellow. A few representative antibiotics in each class are listed. All antibiotics used to treat human infections fall (more...)
Fungal and Protozoan Parasites Have Complex Life Cycles with Multiple Forms
Pathogenic fungi and protozoan parasites are eucaryotes. It is therefore more difficult to find drugs that will kill them without killing the host. Consequently, antifungal and antiparasitic drugs are often less effective and more toxic than antibiotics. A second characteristic of fungal and parasitic infections that makes them difficult to treat is the tendency of the infecting organisms to switch among several different forms during their life cycles. A drug that is effective at killing one form is often ineffective at killing another form, which therefore survives the treatment.
The fungal branch of the eucaryotic kingdom includes both unicellular yeasts (such as Saccharomyces cerevisiae and Schizosaccharomyces pombe) and filamentous, multicellular molds (like those found on moldy fruit or bread). Most of the important pathogenic fungi exhibit dimorphism—the ability to grow in either yeast or mold form. The yeast-to-mold or mold-to-yeast transition is frequently associated with infection. Histoplasma capsulatum, for example, grows as a mold at low temperature in the soil, but it switches to a yeast form when inhaled into the lung, where it can cause the disease histoplasmosis ().
Dimorphism in the pathogenic fungus Histoplasma capsulatum. (A) At low temperature in the soil, Histoplasma grows as a filamentous fungus. (B) After being inhaled into the lung of a mammal, Histoplasma undergoes a morphological switch triggered by the (more...)
Protozoan parasites have more elaborate life cycles than do fungi. These cycles frequently require the services of more than one host. Malaria is the most common protozoal disease, infecting 200–300 million people every year and killing 1–3 million of them. It is caused by four species of Plasmodium, which are transmitted to humans by the bite of the female of any of 60 species of Anopheles mosquito. Plasmodium falciparum—the most intensively studied of the malaria-causing parasites—exists in no fewer than eight distinct forms, and it requires both the human and mosquito hosts to complete its sexual cycle (). Gametes are formed in the bloodstream of infected humans, but they can only fuse to form a zygote in the gut of the mosquito. Three of the Plasmodium forms are highly specialized to invade and replicate in specific tissues—the insect gut lining, the human liver, and the human red blood cell.
The complex life cycle of malaria. (A) The sexual cycle of Plasmodium falciparum requires passage between a human host and an insect host. (B)-(D) Blood smears from people infected with malaria, showing three different forms of the parasite that appear (more...)
Because malaria is so widespread and devastating, it has acted as a strong selective pressure on human populations in areas of the world that harbor the Anopheles mosquito. Sickle cell anemia, for example, is a recessive genetic disorder caused by a point mutation in the gene that encodes the hemoglobin β chain, and it is common in areas of Africa with a high incidence of the most serious form of malaria (caused by Plasmodium falciparum). The malarial parasites grow poorly in red blood cells from either homozygous sickle cell patients or healthy heterozygous carriers, and, as a result, malaria is seldom found among carriers of this mutation. For this reason, malaria has maintained the sickle cell mutation at high frequency in these regions of Africa.
Viruses Exploit Host Cell Machinery for All Aspects of Their Multiplication
Bacteria, fungi, and eucaryotic parasites are cells themselves. Even when they are obligate parasites, they use their own machinery for DNA replication, transcription, and translation, and they provide their own sources of metabolic energy. Viruses, by contrast, are the ultimate hitchhikers, carrying little more than information in the form of nucleic acid. The information is largely replicated, packaged, and preserved by the host cells (). Viruses have a small genome, made up of a single nucleic acid type—either DNA or RNA—which, in either case, may be single-stranded or double-stranded. The genome is packaged in a protein coat, which in some viruses is further enclosed by a lipid envelope.
A simple viral life cycle. The hypothetical virus shown consists of a small double-stranded DNA molecule that codes for only a single viral capsid protein. No known virus is this simple.
Viruses replicate in various ways. In general, replication involves (1) disassembly of the infectious virus particle, (2) replication of the viral genome, (3) synthesis of the viral proteins by the host cell translation machinery, and (4) reassembly of these components into progeny virus particles. A single virus particle (a virion) that infects a single host cell can produce thousands of progeny in the infected cell. Such prodigious viral multiplication is often enough to kill the host cell: the infected cell breaks open (lyses) and thereby allows the progeny viruses access to nearby cells. Many of the clinical manifestations of viral infection reflect this cytolytic effect of the virus. Both the cold sores formed by herpes simplex virus and the lesions caused by the smallpox virus, for example, reflect the killing of the epidermal cells in a local area of infected skin.
Viruses come in a wide variety of shapes and sizes, and, unlike cellular life forms, they cannot be systematically classified by their relatedness into a single phylogenetic tree. Because of their tiny sizes, complete genome sequences have been obtained for nearly all clinically important viruses. Poxviruses are among the largest, up to 450 nm long, which is about the size of some small bacteria. Their genome of double-stranded DNA consists of about 270,000 nucleotide pairs. At the other end of the size scale are parvoviruses, which are less than 20 nm long and have a single-stranded DNA genome of under 5000 nucleotides (). The genetic information in a virus can be carried in a variety of unusual nucleic acid forms ().
Examples of viral morphology. As shown, viruses vary greatly in both size and shape.
Schematic drawings of several types of viral genomes. The smallest viruses contain only a few genes and can have an RNA or a DNA genome. The largest viruses contain hundreds of genes and have a double-stranded DNA genome. The peculiar ends (as well as (more...)
The capsid that encloses the viral genome is made of one or several proteins, arranged in regularly repeating layers and patterns. In enveloped viruses, the capsid itself is enclosed by a lipid bilayer membrane that is acquired in the process of budding from the host cell plasma membrane (). Whereas nonenveloped viruses usually leave an infected cell by lysing it, an enveloped virus can leave the cell by budding, without disrupting the plasma membrane and, therefore, without killing the cell. These viruses can cause chronic infections, and some can help transform an infected cell into a cancer cell.
Acquisition of a viral envelope. (A) Electron micrograph of an animal cell from which six copies of an enveloped virus (Semliki forest virus) are budding. (B) Schematic view of the envelope assembly and budding processes. The lipid bilayer that surrounds (more...)
Despite this variety, all viral genomes encode three types of proteins: proteins for replicating the genome, proteins for packaging the genome and delivering it to more host cells, and proteins that modify the structure or function of the host cell to suit the needs of the virus (). In the second section of this chapter, we focus primarily on this third class of viral proteins.
A map of the HIV genome. This retroviral genome consists of about 9000 nucleotides and contains nine genes, the locations of which are shown in green and red. Three of the genes (green) are common to all retroviruses: gag encodes capsid proteins, env (more...)
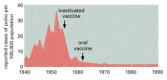
Since most of the critical steps in viral replication are performed by host cell machinery, the identification of effective antiviral drugs is particularly problematic. Whereas the antibiotic tetracycline specifically poisons bacterial ribosomes, for example, it will not be possible to find a drug that specifically poisons viral ribosomes, as viruses use the ribosomes of the host cell to make their proteins. The best strategy for containing viral diseases is to prevent them by vaccination of the potential hosts. Highly successful vaccination programs have effectively eliminated smallpox from the planet, and the eradication of poliomyelitis is imminent ().
Eradication of a viral disease through vaccination. The graph shows number of cases of poliomyelitis reported per year in the United States. The arrows indicate the timing of the introduction of the Salk vaccine (inactivated virus given by injection) (more...)
Prions Are Infectious Proteins
All information in biological systems is encoded by structure. We are used to thinking of biological information in the form of nucleic acid sequences (as in our description of viral genomes), but the sequence itself is a shorthand code for describing nucleic acid structure. The replication and expression of the information encoded in DNA and RNA are strictly dependent on the structure of these nucleic acids and their interactions with other macromolecules. The propagation of genetic information primarily requires that the information be stored in a structure that can be duplicated from unstructured precursors. Nucleic acid sequences are the simplest and most robust solution that organisms have found to the problem of faithful structural replication.
Nucleic acids are not the only solution, however. Prions are infectious agents that are replicated in the host by copying an aberrant protein structure. They can occur in yeasts, and they cause various neurodegenerative diseases in mammals. The most well-known infection caused by prions is bovine spongiform encephalopathy (BSE, or mad cow disease), which occasionally spreads to humans who eat infected parts of the cow (). Isolation of the infectious prions that cause the disease scrapie in sheep, followed by years of painstaking laboratory characterization of scrapie-infected mice, eventually established that the protein itself is infectious.
Neural degeneration in a prion infection. This micrograph shows a slice from the brain of a person who died of kuru. Kuru is a human prion disease, very similar to BSE, that was spread from one person to another by ritual mortuary practices in New Guinea. (more...)
Intriguingly, the infectious prion protein is made by the host, and its amino acid sequence is identical to a normal host protein. Moreover, the prion and normal forms of the protein are indistinguishable in their posttranslational modifications. The only difference between them appears to be in their folded three-dimensional structure. The misfolded prion protein tends to aggregate, and it has the remarkable capacity to cause the normal protein to adopt its misfolded prion conformation and thereby to become infectious (see ). This ability of the prion to convert the normal host protein to misfolded prion protein is equivalent to the prion's having replicated itself in the host. If eaten by another susceptible host, these newly-misfolded prions can transmit the infection.
It is not known how normal proteins are usually able to find the single, correct, folded conformation, among the billions of other possibilities, without becoming stuck in dead-end intermediates (discussed in Chapters 3 and 6). Prions are a good example of how protein folding can go dangerously wrong. But, why are the prion diseases so uncommon? What are the constraints that determine whether a misfolded protein will behave like a prion, or simply get refolded or degraded by the cell that made it? We do not yet have answers to these questions, and the study of prions remains an area of intense research.
Summary
Infectious diseases are caused by pathogens, which include bacteria, fungi, protozoa, worms, viruses, and even infectious proteins called prions. Pathogens of all classes must have mechanisms for entering their host and for evading immediate destruction by the host immune system. Most bacteria are not pathogenic. Those that are contain specific virulence genes that mediate interactions with the host, eliciting particular responses from the host cells that promote the replication and spread of the pathogen. Pathogenic fungi, protozoa, and other eucaryotic parasites typically pass through several different forms during the course of infection; the ability to switch among these forms is usually required for the parasites to be able to survive in a host and cause disease. In some cases, such as malaria, parasites must pass sequentially through several host species to complete their life cycles. Unlike bacteria and eucaryotic parasites, viruses have no metabolism of their own and no intrinsic ability to produce the proteins encoded by their DNA or RNA genomes. They rely entirely on subverting the machinery of the host cell to produce their proteins and to replicate their genomes. Prions, the smallest and simplest infectious agents, contain no nucleic acid; instead, they are rare, aberrantly folded proteins that happen to catalyze the misfolding of proteins in the host that share their primary amino acid sequence.