By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Copyright © 2000, Sinauer
Associates.
Bookshelf ID: NBK10008

An official website of the United States government

NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
Gilbert SF. Developmental Biology. 6th edition. Sunderland (MA): Sinauer Associates; 2000.

Oogenesis—the differentiation of the ovum—differs from spermatogenesis in several ways. Whereas the gamete formed by spermatogenesis is essentially a motile nucleus, the gamete formed by oogenesis contains all the materials needed to initiate and maintain metabolism and development. Therefore, in addition to forming a haploid nucleus, oogenesis also builds up a store of cytoplasmic enzymes, mRNAs, organelles, and metabolic substrates. While the sperm becomes differentiated for motility, the egg develops a remarkably complex cytoplasm.
The mechanisms of oogenesis vary among species more than those of spermatogenesis. This difference should not be surprising, since patterns of reproduction vary so greatly among species. In some species, such as sea urchins and frogs, the female routinely produces hundreds or thousands of eggs at a time, whereas in other species, such as humans and most mammals, only a few eggs are produced during the lifetime of an individual. In those species that produce thousands of ova, the oogonia are self-renewing stem cells that endure for the lifetime of the organism. In those species that produce fewer eggs, the oogonia divide to form a limited number of egg precursor cells. In the human embryo, the thousand or so oogonia divide rapidly from the second to the seventh month of gestation to form roughly 7 million germ cells (Figure 19.19). After the seventh month of embryonic development, however, the number of germ cells drops precipitously. Most oogonia die during this period, while the remaining oogonia enter the first meiotic division (Pinkerton et al. 1961). These latter cells, called the primary oocytes, progress through the first meiotic prophase until the diplotene stage, at which point they are maintained until puberty. With the onset of adolescence, groups of oocytes periodically resume meiosis. Thus, in the human female, the first part of meiosis begins in the embryo, and the signal to resume meiosis is not given until roughly 12 years later. In fact, some oocytes are maintained in meiotic prophase for nearly 50 years. As Figure 19.19 indicates, primary oocytes continue to die even after birth. Of the millions of primary oocytes present at birth, only about 400 mature during a woman's lifetime.

Changes in the number of germ cells in the human ovary over the life span. (After Baker 1970.)
Oogenic meiosis also differs from spermatogenic meiosis in its placement of the metaphase plate. When the primary oocyte divides, its nucleus, called the germinal vesicle, breaks down, and the metaphase spindle migrates to the periphery of the cell. At telophase, one of the two daughter cells contains hardly any cytoplasm, whereas the other cell has nearly the entire volume of cellular constituents (Figure 19.20). The smaller cell is called the first polar body, and the larger cell is referred to as the secondary oocyte. During the second division of meiosis, a similar unequal cytokinesis takes place. Most of the cytoplasm is retained by the mature egg (ovum), and a second polar body receives little more than a haploid nucleus. Thus, oogenic meiosis conserves the volume of oocyte cytoplasm in a single cell rather than splitting it equally among four progeny.

Polar body formation in the oocyte of the whitefish Coregonus. (A) Anaphase of first meiotic division, showing the first polar body pinching off with its chromosomes. (B) Metaphase (within the oocyte, arrow) of the second meiotic division, with the first (more...)
In a few species of animals, meiosis is severely modified such that the resulting gamete is diploid and need not be fertilized to develop. Such animals are said to be parthenogenetic (Greek, “virgin birth”). In the fly Drosophila mangabeirai, one of the polar bodies acts as a sperm and “fertilizes” the oocyte after the second meiotic division. In other insects (such as Moraba virgo) and in the lizard Cnemidophorus uniparens, the oogonia double their chromosome number before meiosis, so that the halving of the chromosomes restores the diploid number. The germ cells of the grasshopper Pycnoscelus surinamensis dispense with meiosis altogether, forming diploid ova by two mitotic divisions (Swanson et al. 1981). All of these species consist entirely of females. In other species, haploid parthenogenesis is widely used not only as a means of reproduction, but also as a mechanism of sex determination. In the Hymenoptera (bees, wasps, and ants), unfertilized eggs develop into males, whereas fertilized eggs, being diploid, develop into females. The haploid males are able to produce sperm by abandoning the first meiotic division, thereby forming two sperm cells through second meiosis.
The egg is responsible for initiating and directing development, and in some species (as seen above), fertilization is not even necessary. The accumulated material in the oocyte cytoplasm includes energy sources and energy-producing organelles (the yolk and mitochondria); the enzymes and precursors for DNA, RNA, and protein syntheses; stored messenger RNAs; structural proteins; and morphogenetic regulatory factors that control early embryogenesis. A partial catalogue of the materials stored in the oocyte cytoplasm is shown in Table 19.2, while a partial list of stored mRNAs is shown in Table 5.3. Most of this accumulation takes place during meiotic prophase I, and this stage is often subdivided into two phases, previtellogenesis (Greek, “before yolk formation”) and vitellogenesis.
Cellular components stored in the mature oocyte of Xenopus laevis.
The eggs of fishes and amphibians are derived from an oogonial stem cell population that can generate a new cohort of oocytes each year. In the frog Rana pipiens, oogenesis takes 3 years. During the first 2 years, the oocyte increases its size very gradually. During the third year, however, the rapid accumulation of yolk in the oocyte causes the egg to swell to its characteristically large size (Figure 19.21). Eggs mature in yearly batches, with the first cohort maturing shortly after metamorphosis; the next group matures a year later.

Growth of oocytes in the frog. During the first 3 years of life, three cohorts of oocytes are produced. The drawings follow the growth of the first-generation oocytes. (After Grant 1953.)
Vitellogenesis occurs when the oocyte reaches the diplotene stage of meiotic prophase. Yolk is not a single substance, but a mixture of materials used for embryonic nutrition. The major yolk component in frog eggs is a 470-kDa protein called vitellogenin. It is not made in the frog oocyte (as are the major yolk proteins of organisms such as annelids and crayfishes), but is synthesized in the liver and carried by the bloodstream to the ovary (Flickinger and Rounds 1956). This large protein passes between the follicle cells of the ovary, and is incorporated into the oocyte by micropinocytosis, the pinching off of membrane-bounded vesicles at the bases of microvilli (Dumont 1978). In the mature oocyte, vitellogenin is split into two smaller proteins: the heavily phosphorylatedphosvitin and the lipoprotein lipovitellin. These two proteins are packaged together into membrane-bounded yolk platelets (Figure 19.22A). Glycogen granules and lipochondrial inclusions store the carbohydrate and lipid components of the yolk, respectively.
Yolk distribution in Xenopus. (A) An amphibian yolk platelet. (B-E) Establishment of animal-vegetal distribution of yolk platelets in the Xenopus oocyte. (B) In the late stage III (600 μm) oocyte, yolk platelets enter the cell equally at all points (more...)
Most eggs are highly asymmetrical, and it is during oogenesis that the animal-vegetal axis of the egg is specified. Danilchik and Gerhart (1987) have shown that although the concentration of yolk increases nearly tenfold as one moves from the animal to the vegetal poles of the mature Xenopus egg, vitellogenin uptake is uniform around the surface of the oocyte. What varies is its movement within the oocyte, and this depends on where the yolk proteins enter. When yolk platelets are formed in the future animal hemisphere, they move inward toward the center of the cell. Vegetal yolk platelets, however, do not actively move, but remain at the periphery of the cell for long periods of time, enlarging as they stay there. They are slowly displaced from the cortex as new yolk platelets come in from the surface. As a result of this differential intracellular transport, the amount of yolk steadily increases in the vegetal hemisphere, until the vegetal half of a mature Xenopus oocyte contains nearly 75% of the yolk (Figure 19.22B-E). The mechanism of this translocation remains unknown.
As the yolk is being deposited, the organelles also become arranged asymmetrically. The cortical granules begin to form from the Golgi apparatus; they are originally scattered randomly through the oocyte cytoplasm, but later migrate to the periphery of the cell. The mitochondria replicate at this time, dividing to form millions of mitochondria that will be apportioned to the different blastomeres during cleavage. (In Xenopus, new mitochondria will not be formed until after gastrulation is initiated.) As vitellogenesis nears an end, the oocyte cytoplasm becomes stratified. The cortical granules, mitochondria, and pigment granules are found at the periphery of the cell, within the actin-rich oocyte cortex. Within the inner cytoplasm, distinct gradients emerge. While the yolk platelets become more heavily concentrated at the vegetal pole of the oocyte, the glycogen granules, ribosomes, lipid vesicles, and endoplasmic reticulum are found toward the animal pole. Even specific mRNAs stored in the cytoplasm become localized to certain regions of the oocyte.
While the precise mechanisms for establishing these gradients remain unknown, studies using inhibitors have shown that the cytoskeleton is critically important in localizing specific RNAs and morphogenetic factors. There seem to be two pathways for gettting mRNAs into the vegetal cortex (Forristall et al. 1995; Kloc and Etkin 1995, Kloc et al. 1998). The first pathway moves messages such as those encoding the Vg1 protein, which are initially present throughout the oocyte, into the vegetal cortex in a two-step process (Yisraeli et al. 1990). In the first phase, microtubules are needed to bring Vg1 mRNA into the vegetal hemisphere. In the second phase, microfilaments are responsible for anchoring the Vg1 message to the cortex. The portion of the Vg1 mRNA that binds to these cytoskeletal elements resides in its 3´ untranslated region. When a specific 340-base sequence from the Vg1 3´ UTR is placed onto a β-globin message, that β-globin mRNA is similarly localized to the vegetal cortex (see Chapter 5; Mowry and Melton 1992). Other mRNAs, including germ plasm mRNAS such as Xlsirt and Xcat2, leave the germinal vesicle and join the mitochondrial “cloud” located at the vegetal pole of the nucleus. These messages are compartmentalized into clusters associated with the germ plasm and transported to the vegetal cortex in a manner that appears to be independent of the cytoskeleton (Figure 19.23; Kloc et al. 1996). This mechanism is known as the Metro (message transport organizer) pathway.

Schematic representations of the two pathways for localizing mRNAs to the vegetal region of the Xenopus oocyte. The respective mRNAs are shown in yellow. In the cytoskeletal (Vg1) pathway, messages are first seen throughout the egg, but they are translocated (more...)
19.9 Hormonal control of yolk production. Vitellogenesis in amphibians is mediated primarily by estrogen. Estrogen instructs the liver to express and secrete vitellogenin, and this protein is absorbed from the blood by the young oocyte. http://www.devbio.com/chap19/link1909.shtml
19.10 Transporting the Vg1 mRNA. The Vera protein specifically binds to the 3´ UTR of the Vg1 message. Vera may link Vg1 mRNA to a set of endoplasmic reticulum vesicles that are translocated to the vegetal cortex. http://www.devbio.com/chap19/link1910.shtml
19.11 Establishment of egg polarity. In several species, the developing oocyte is a flagellated cell whose flagellum marks the future animal pole of the egg. This flagellum is lost during oogenesis. http://www.devbio.com/chap19/link1911.shtml
Amphibian oocytes can remain for years in the diplotene stage of meiotic prophase. This state resembles the G2 phase of the cell division cycle (see Chapter 8). Resumption of meiosis in the amphibian primary oocyte requires progesterone. This hormone is secreted by the follicle cells in response to gonadotropic hormones secreted by the pituitary gland. Within 6 hours of progesterone stimulation, germinal vesicle breakdown (GVBD) occurs, the microvilli retract, the nucleoli disintegrate, and the chromosomes contract and migrate to the animal pole to begin division. Soon afterward, the first meiotic division occurs, and the mature ovum is released from the ovary by a process called ovulation. The ovulated egg is in second meiotic metaphase when it is released (Figure 19.24).

Schematic representation of Xenopus oocyte maturation, showing the regulation of meiotic cell division by progesterone and fertilization. Oocyte maturation is blocked in the diplotene stage of first meiotic prophase by the lack of active MPF. Progesterone (more...)
How does progesterone enable the egg to break its dormancy and resume meiosis? To understand the mechanisms by which this activation is accomplished, it is necessary to briefly review the model for early blastomere division (see Chapter 8). Entry into the mitotic (M) phase of the cell cycle (in both meiosis and mitosis) is regulated by mitosis-promoting factor, or MPF (originally called “maturation-promoting factor” after its meiotic function). MPF contains two subunits, cyclin B and the p34cdc2 protein. The p34 protein is a cyclin-dependent-kinase—its activity is dependent upon the presence of cyclin. Since all the components of MPF are present in the amphibian oocyte, it is generally thought that progesterone somehow converts a pre-MPF complex into active MPF.
The mediator of the progesterone signal is the c-mos protein. Progesterone reinitiates meiosis by causing the egg to polyadenylate the maternal c-mos mRNA that has been stored in its cytoplasm (Sagata et al. 1988, 1989; Sheets et al. 1995). This message is translated into a 39-kDa phosphoprotein, known as c-mos. This protein is detectable only during oocyte maturation and is destroyed quickly upon fertilization. Yet during its brief lifetime, it plays a major role in releasing the egg from its dormancy. If the translation of c-mos is inhibited (by injecting c-mos antisense mRNA into the oocyte), germinal vesicle breakdown and the resumption of oocyte maturation do not occur. The c-mos protein activates a phosphorylation cascade that phosphorylates and activates the p34 subunit of MPF (Ferrell and Machleder 1998; Ferrell 1999). The active MPF allows the germinal vesicle to break down and the chromosomes to divide.
However, the chromosomes then encounter a second block. MPF can take the chromosomes through only the first meiotic division and the prophase of the second meiotic division. The oocyte is arrested again in the metaphase of the second meiotic division. This metaphase block is caused by the combined actions of c-mos and another protein, cyclin-dependent kinase 2 (cdk2; Gabrielli et al. 1993). These two proteins are subunits of cytostatic factor (CSF), which is found in mature frog eggs, and which can block cell cycles in metaphase (Matsui 1974). It is thought that CSF prevents the degradation of cyclin (Figure 19.24).
The metaphase block is broken by fertilization. Evidence suggests that the calcium ion flux attending fertilization enables the calcium-binding protein calmodulin to become active. Calmodulin, in turn, can activate two enzymes that inactivate CSF: calmodulin-dependent protein kinase II, which inactivates p34, and calpain II, a calcium-dependent protease that degrades c-mos (Watanabe et al. 1989; Lorca et al. 1993). Without CSF, cyclin can be degraded, and the meiotic division can be completed.
In most animals (insects being a major exception), the growing oocyte is active in transcribing genes whose products are (1) necessary for cell metabolism, (2) necessary for oocyte-specific processes, or (3) needed for early development before the zygote-derived nuclei begins to function. In mice, for instance, the growing diplotene oocyte is actively transcribing the genes for zona pellucida proteins ZP1, ZP2, and ZP3. Moreover, these genes are transcribed only in the oocyte and not in any other cell (Figure 19.25; Roller et al. 1989; Lira et al. 1990; Epifano et al. 1995).

Expression of the ZP3 gene in the developing mouse oocyte. (A) Northern blot of ZP3 mRNA accumulation in embryonic mouse tissues. A radioactive probe to the ZP3 message found it expressed only in the ovary, and specifically in the oocytes. (B-C) When (more...)
The amphibian oocyte has certain periods of very active RNA synthesis. During the diplotene stage, certain chromosomes stretch out large loops of DNA, causing the chromosome to resemble a lampbrush (a handy instrument for cleaning test tubes in the days before microfuges). These lamp brush chromosomes (Figure 19.26) can be revealed as the sites of RNA synthesis by in situ hybridization. Oocyte chromosomes can be incubated with a radioactive RNA probe, and autoradiography used to visualize the precise location where the gene is being transcribed. Figure 19.26B shows diplotene chromosome I of the newt Triturus cristatus after incubation with radioactive histone mRNA. It is obvious that a histone gene (or set of histone genes) is located on one of these loops of the lampbrush chromosome (Old et al. 1977). Electron micrographs of gene transcripts from lampbrush chromosomes also enable one to see chains of mRNA coming off each gene as it is transcribed (Hill and MacGregor 1980).

Amphibian lampbrush chromosomes are active in the diplotene germinal vesicle during first meiotic prophase. (A) A lampbrush chromosome of the salamander Notophthalmus viridescens. Extended DNA (white) loops out and is transcribed into RNA (red). (B) Localization (more...)
In addition to mRNA synthesis, the patterns of ribosomal RNA and transfer RNA transcription are also regulated during oogenesis. Figure 19.27A shows the course of rRNA and tRNA synthesis during Xenopus oogenesis. Transcription appears to begin in early (stage I, 25–40 μm) oocytes, during the diplotene stage of meiosis. At this time, all the rRNAs and tRNAs needed for protein synthesis until the mid-blastula stage are made, and all the maternal mRNAs needed for early development are transcribed. This stage lasts for months in Xenopus. The rate of ribosomal RNA production is prodigious. The Xenopus oocyte genome has over 1800 genes encoding 18S and 28S rRNA (the two large RNAs that form the ribosomes), and these genes are selectively amplified such that there are over 500,000 genes making these ribosomal RNAs (Figure 19.27B; Brown and Dawid 1968). After reaching a certain size, the chromosomes of the mature (stage VI) oocyte condense, and the genes are no longer transcribed. This “mature oocyte” condition can also last for months. Upon hormonal stimulation, the oocyte completes its first meiotic division and is ovulated. The mRNAs stored by the oocyte now join with the ribosomes to initiate protein synthesis. Within hours, the second meiotic division has begun, and the secondary oocyte has been fertilized. The embryo's genes do not begin active transcription until the mid-blastula transition (Davidson 1986).

Ribosomal RNA production in Xenopus oocytes. (A) Relative rates of DNA, tRNA, and rRNA synthesis in amphibian oogenesis during the last 3 months before ovulation. (B) The transcription of the RNA precursor of the 28S, 18S, and 5.8S ribosomal RNAs. These (more...)
As we have seen in Chapter 5, the oocytes of several species make two classes of mRNAs—those for immediate use in the oocyte and those that are stored for use during early development. In sea urchins, the translation of stored maternal messages is initiated by fertilization, while in frogs, the signal for such translation is initiated by progesterone as the egg is about to be ovulated. One of the results of the MPF activity induced by progesterone may be the phosphorylation of proteins on the 3´ UTR of stored oocyte mRNAs. The phosphorylation of these factors is associated with the lengthening of the poly(A) tails in the stored messages and the translation of the stored mRNAs (Paris et al. 1991).
19.12 Synthesizing oocyte ribosomes. Ribosomes are almost a “differentiated product” of the oocyte, and the Xenopus oocyte contains 20,000 times as many ribosomes as somatic cells do. Gene repetition and gene amplification are both used to transcribe these enormous amounts of rRNA. http://www.devbio.com/chap19/link1912.shtml
There are several types of oogenesis in insects, but most studies have focused on those insects, such as Drosophila and moths, that undergo meroistic oogenesis, in which cytoplasmic connections remain between the cells produced by the oogonium. In Drosophila, each oogonium divides four times to produce a clone of 16 cells connected to each other by ring canals. The production of these interconnected cells, called cystocytes, involves a highly ordered array of cell divisions (Figure 19.28). Only those two cells having four interconnections are capable of developing into oocytes, and of those two, only one becomes the egg. The other begins meiosis but does not complete it. Thus, only one of the 16 cystocytes can become an ovum. All the other cells become nurse cells. As it turns out, the cell destined to become the oocyte is that cell residing at the most posterior tip of the egg chamber, or ovariole, that encloses the 16-cell clone. However, since the nurse cells are connected to the oocyte by cytoplasmic bridges, the entire complex can be seen as one egg-producing unit.

The formation of 16 interconnected cystocytes in Drosophila. (A) Diagram of adult ovariole, showing sequence of oogenesis as younger germinal cysts mature within the ovariole. (B) Section through center of an ovariole, showing the maturation of the oocyte. (more...)
The meroistic ovary confronts us with some interesting problems. If all the cystocytes are connected so that proteins and RNAs shuttle freely among them, why should they have different developmental fates? Why should one cell become the oocyte while the others become “RNA-synthesizing factories,” sending mRNAs, ribosomes, and even centrioles into the oocyte? Why is the flow of protein and RNA in one direction only? As the cystocytes divide, a large, spectrin-rich structure called the fusome forms and spans the ring canals between the cells (Figure 19.28C). It is constructed asymmetrically, as it always grows from the spindle pole that remained in one of the cells after the first division (Lin and Spradling 1995; de Cuevas and Spradling 1998). The cell that retained the greater part of the fusome during the first division becomes the oocyte. It is not yet known if the fusome contains oogenic determinants, or if it directs the traffic of materials into this particular cell.
Once the patterns of transport are established, the cytoskeleton becomes actively involved in transporting mRNAs from the nurse cells into the oocyte cytoplasm (Cooley and Theurkauf 1994). The microtubular array is critical for oocyte determination. If this lattice is disrupted (either chemically or by mutations such as bicaudal-D or egalitarian), gene products are transmitted in all directions, and all 16 cells differentiate into nurse cells (Gutzeit 1986; Theurkauf et al. 1992, 1993; Spradling 1993). It is possible that some compounds transported from the nurse cells into the oocyte become associated with transport proteins, such as kinesin, that would enable them to travel along the tracks of microtubules extending through the ring canals (Theurkauf et al. 1992; Sun and Wyman 1993). Actin may become important for maintaining this polarity during later stages of oogenesis. Mutations that prevent actin microfilaments from lining the ring canals prevent the transport of mRNAs from nurse cells to oocyte, and disruption of the actin microfilaments randomizes the distribution of mRNA (Cooley et al. 1992; Watson et al. 1993). Thus, the cytoskeleton appears to control the movement of organelles and RNAs between nurse cells and oocyte such that developmental cues are exchanged only in the appropriate direction.
The oocytes of meroistic insects do not pass through a transcriptionally active stage, nor do they have lampbrush chromosomes. Rather, RNA synthesis is largely confined to the nurse cells, and the RNA made by those cells is actively transported into the oocyte cytoplasm. This can be seen in Figure 9.36A. Oogenesis takes place in only 12 days, so the nurse cells are very metabolically active during this time. They are aided in their transcriptional efficiency by becoming polytene. Instead of having two copies of each chromosome, they replicate their chromosomes until they have produced 512 copies. The 15 nurse cells pass ribosomal and messenger RNAs as well as proteins into the oocyte cytoplasm, and entire ribosomes may be transported as well. The mRNAs do not associate with polysomes, which suggests that they are not immediately active in protein synthesis (Paglia et al. 1976; Telfer et al. 1981).
Ovulation of the mammalian egg follows one of two basic patterns, depending on the species. One type of ovulation is stimulated by the act of copulation. Physical stimulation of the cervix triggers the release of gonadotropins from the pituitary. These gonadotropins signal the egg to resume meiosis and initiate the events that will expel it from the ovary. This mechanism ensures that most copulations will result in fertilized ova, and animals that utilize this method of ovulation—rabbits and minks—have a reputation for procreative success.
Most mammals, however, have a periodic ovulation pattern, in which the female ovulates only at specific times of the year. This ovulatory time is called estrus (or its English equivalent, “heat”). In these cases, environmental cues, most notably the amount and type of light during the day, stimulate the hypothalamus to release gonadotropin-releasing factor. This factor stimulates the pituitary to release its gonadotropins—follicle-stimulating hormone (FSH) and luteinizing hormone (LH)—which cause the follicle cells to proliferate and secrete estrogen. The estrogen enters certain neurons and evokes the pattern of mating behavior characteristic of the species. The gonadotropins also stimulate follicular growth and initiate ovulation. Thus, estrus and ovulation occur close together.
Humans have a variation on the theme of periodic ovulation. Although human females have cyclical ovulation (averaging about once every 29.5 days) and no definitive yearly estrus, most of human reproductive physiology is shared with other primates. The characteristic primate periodicity in maturing and releasing ova is called the menstrual cycle because it entails the periodic shedding of blood and endothelial tissue from the uterus at monthly intervals.* The menstrual cycle represents the integration of three very different cycles: (1) the ovarian cycle, the function of which is to mature and release an oocyte, (2) the uterine cycle, the function of which is to provide the appropriate environment for the developing blastocyst, and (3) the cervical cycle, the function of which is to allow sperm to enter the female reproductive tract only at the appropriate time. These three functions are integrated through the hormones of the pituitary, hypothalamus, and ovary.
Oogenesis in mammals. The development of the mammalian ovum and its remarkable growth during its primary oocyte stage is the subject of photographs and QuickTime movies of histological sections through a mammalian ovary.[Click on Gametogenesis]
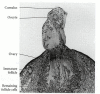
The majority of the oocytes within the adult human ovary are maintained in the prolonged diplotene stage of the first meiotic prophase (often referred to as the dictyate state). Each oocyte is enveloped by a primordial follicle consisting of a single layer of epithelial granulosa cells and a less organized layer of mesenchymal thecal cells (Figure 19.29). Periodically, a group of primordial follicles enters a stage of follicular growth. During this time, the oocyte undergoes a 500-fold increase in volume (corresponding to an increase in oocyte diameter from 10 μm in a primordial follicle to 80 μm in a fully developed follicle). Concomitant with oocyte growth is an increase in the number of follicular granulosa cells, which form concentric layers around the oocyte. This proliferation of granulosa cells is mediated by a paracrine factor, GDF9, a member of the TGF-β family (Dong et al. 1996). Throughout this growth period, the oocyte remains in the dictyate stage. The fully grown follicle thus contains a large oocyte surrounded by several layers of granulosa cells. The innermost of these cells will stay with the ovulated egg, forming the cumulus, which surrounds the egg in the oviduct. In addition, during the growth of the follicle, an antrum (cavity) forms, which becomes filled with a complex mixture of proteins, hormones, and other molecules. Just as the maturing oocyte synthesizes paracrine factors that allow the follicle cells to proliferate, the follicle cells secrete growth and differentiation factors (TGF-β2, VEGF, leptin, FGF2) that allow the oocyte to grow and which bring blood vessels into the follicular region (Antczak et al. 1997).

The ovarian follicle of mammals. (A) Maturation of the ovarian follicle. When mature, it is often called a Graafian follicle. (B) Scanning electron micrograph of a mature follicle in the rat. The oocyte (center) is surrounded by the smaller granulosa (more...)
At any given time, a small group of follicles is maturing. However, after progressing to a certain stage, most oocytes and their follicles die. To survive, the follicle must be exposed to gonadotropic hormones and, “catching the wave” at the right time, must ride it until it peaks. Thus, for oocyte maturation to occur, the follicle needs to be at a certain stage of development when the waves of gonadotropin arise.
The first day of vaginal bleeding is considered to be day 1 of the menstrual cycle (Figure 19.30). This bleeding represents the sloughing off of endometrial tissue and blood vessels that would have aided the implantation of the blastocyst. In the first part of the cycle (called the proliferative or follicular phase), the pituitary starts secreting increasingly large amounts of FSH. Any maturing follicles that have reached a certain stage of development respond to this hormone with further growth and cellular proliferation. FSH also induces the formation of LH receptors on the granulosa cells. Shortly after this period of initial follicle growth, the pituitary begins secreting LH. In response to LH, the dictyate meiotic block is broken. The nuclear membranes of competent oocytes break down, and the chromosomes assemble to undergo the first meiotic division. One set of chromosomes is kept inside the oocyte, and the other ends up in the small polar body. Both are encased by the zona pellucida, which has been synthesized by the growing oocyte. It is at this stage that the egg will be ovulated.
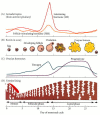
The human menstrual cycle. The coordination of (B) ovarian and (D) uterine cycles is controlled by (A) the pituitary and (C) the ovarian hormones. During the follicular phase, the egg matures within the follicle, and the uterine lining is prepared to (more...)
The two gonadotropins, acting together, cause the follicle cells to produce increasing amounts of estrogen, which has at least five major activities in regulating the further progression of the menstrual cycle:
It causes the uterine endometrium to begin its proliferation and to become enriched with blood vessels.
It causes the cervical mucus to thin, thereby permitting sperm to enter the inner portions of the reproductive tract.
It causes an increase in the number of FSH receptors on the granulosa cells of the mature follicles (Kammerman and Ross 1975) while causing the pituitary to lower its FSH production. It also stimulates the granulosa cells to secrete the peptide hormone inhibin, which also suppresses pituitary FSH secretion (Rivier et al. 1986; Woodruff et al. 1988).
At low concentrations, it inhibits LH production, but at high concentrations, it stimulates it.
At very high concentrations and over long durations, estrogen interacts with the hypothalamus, causing it to secrete gonadotropin-releasing factor.
As estrogen levels increase as a result of follicular production, FSH levels decline. LH levels, however, continue to rise as more estrogen is secreted. As estrogens continue to be made (days 7–10), the granulosa cells continue to grow. Starting on day 10, estrogen secretion rises sharply. This rise is followed at midcycle by an enormous surge of LH and a smaller burst of FSH. Experiments with female monkeys have shown that exposure of the hypothalamus to greater than 200 pg of estrogen per milliliter of blood for more than 50 hours results in the hypothalamic secretion of gonadotropin-releasing factor. This factor causes the subsequent release of FSH and LH from the pituitary. Within 10 to 12 hours after the gonadotropin peak, the egg is ovulated (Figure 19.31; Garcia et al. 1981).
Ovulation in the rabbit. The ovary of a living, anesthetized rabbit was exposed and observed. When the follicle started to ovulate, the ovary was removed, fixed, and stained. (Photograph courtesy of R. J. Blandau.)
Although the detailed mechanism of ovulation is not yet known, the physical expulsion of the mature oocyte from the follicle appears to be due to an LH-induced increase in collagenase, plasminogen activator, and prostaglandin within the follicle (Lemaire et al. 1973). The mRNA for plasminogen activator has been dormant in the oocyte cytoplasm. LH causes this message to be polyadenylated and translated into this powerful protease (Huarte et al. 1987). Prostaglandins may cause localized contractions in the smooth muscles in the ovary and may also increase the flow of water from the ovarian capillaries, increasing fluid pressure in the antrum (Diaz-Infante et al. 1974; Koos and Clark 1982). If ovarian prostaglandin synthesis is inhibited, ovulation does not take place. In addition, collagenase and the plasminogen activator protease loosen and digest the extracellular matrix of the follicle (Beers et al. 1975; Downs and Longo 1983). The result of LH, then, is increased follicular pressure coupled with the degradation of the follicle wall. A hole is digested through which the ovum can burst.
Following ovulation, the luteal phase of the menstrual cycle begins. The remaining cells of the ruptured follicle, under the continued influence of LH, become the corpus luteum. (They are able to respond to this LH because the surge in FSH stimulates them to develop even more LH receptors.) The corpus luteum secretes some estrogen, but its predominant secretion is progesterone. This steroid hormone circulates to the uterus, where it completes the job of preparing the uterine tissue for blastocyst implantation, stimulating the growth of the uterine wall and its blood vessels. Blocking the progesterone receptor with the synthetic steroid mifepristone (RU486) stops the uterine wall from thickening and prevents the implantation of a blastocyst† (Couzinet et al. 1986; Greb et al. 1999). Progesterone also inhibits the production of FSH, thereby preventing the maturation of any more follicles and ova. (For this reason, a combination of estrogen and progesterone has been used in birth control pills. The growth and maturation of new ova are prevented as long as FSH is inhibited.)
If the ovum is not fertilized, the corpus luteum degenerates, progesterone secretion ceases, and the uterine wall is sloughed off. With the decline in serum progesterone levels, the pituitary secretes FSH again, and the cycle is renewed. However, if fertilization occurs, the trophoblast secretes a new hormone, luteotropin, which causes the corpus luteum to remain active and serum progesterone levels to remain high. Thus, the menstrual cycle enables the periodic maturation and ovulation of human ova and allows the uterus to periodically develop into an organ capable of nurturing a developing organism for 9 months.
19.13 The reinitiation of mammalian meiosis. The hormone-mediated disruption of communication between the oocyte and its surrounding follicle cells may critical in the resumption of meiosis in female mammals. http://www.devbio.com/chap19/link1913.shtml
The egg and the sperm will both die if they do not meet. We are now back where we began: the stage is set for fertilization to take place. As F. R. Lillie recognized in 1919, “The elements that unite are single cells, each on the point of death; but by their union a rejuvenated individual is formed, which constitutes a link in the eternal process of Life.”
The periodic shedding of the uterine lining is a controversial topic. Some scientists speculate that menstruation is an active process, with adaptive significance in evolution. Profet (1993) proposed that menstruation is a crucial immunological adaptation, protecting the uterus against infections from semen or other environmental agents. Strassmann (1996) suggests that the cyclicity of the endometrium is an energy-saving adaptation that is important in times of poor nutrition. Vaginal bleeding would be a side effect of this adaptive process. Finn (1998) claims that menstruation has no adaptive value and is necessitated by the immunological crises that are a consequence of bringing two genetically dissimilar organisms together in the uterus. Martin (1992) points out that it might even be wrong to think of there being a single function of menstruation, and that its roles might change during a woman's life cycle.
RU486 is thought to compete for the progesterone receptor inside the nucleus. RU486 can bind to the progesterone site in the receptor, and the receptor-RU486 complex appears to form heterodimers with the normal progesterone-carrying progesterone receptor. When this RU486-progesterone complex binds to the progesterone-responsive enhancer elements on the DNA, transcription from this site is inhibited (Vegeto et al. 1992; Spitz and Bardin 1993). In Europe, RU486 has become a widely used alternative to surgical abortion (Palka 1989; Maurice 1991).
By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
Your browsing activity is empty.
Activity recording is turned off.
See more...